Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Subjects:
Marine & Freshwater Biology
Ecosystem services (ES) are defined as the benefits that humans obtain from ecological systems. These include services such as food and fresh water and climate regulation, among others that make human life possible. This concept is established as a “policy advocacy tool” since it helps with management practices.
- biodiversity
- Chile
- Lessonia
- Macrocystis
- Peru
1. The Forest-Forming Species of the Humboldt Current System
In the HCS, kelps commonly occur in subtidal rocky reefs except in the most sheltered or turbid locations, and from the lower shores to depths near 30 m [55,56,57]. However, kelp forests composed of Lessonia and Macrocystis have been found in deeper waters (>30 m) [58]. It has been observed that M. pyrifera usually forms an upper canopy, while L. trabeculata, a lower one [45], playing different roles as habitat-structuring species when occurring in the same forest. The distribution patterns of the four kelp species addressed in this review are shown in Figure 1.

Figure 1. Distribution patterns of kelp species forming marine forests in the Humboldt Current System: Macrocysstis pyrifera (brown), Lessonia trabeculata (white), L. berteroana (yellow), and L. spicata (orange).
The kelp forest metacommunities are exposed to different regimes, such as permanent (Peru) or seasonal (Chile) upwelling patterns. The forests located at the northern extreme of the HCS are directly influenced by events associated with the El Niño Southern Oscillation (ENSO), such as the “El Niño Costero” event, which changes the biogeochemical properties of the northern HCS that reduces the availability of nitrates on the upper layer of coastal waters [59] and increases the temperature. On the other hand, cold and eutrophic sub-Antarctic waters predominantly influence the forests in the southern extreme. Although species distribution patterns have been studied for decades, it is still a challenge to monitor the associated assemblages or the ecosystem dynamic at a wide scale in the HCS [60,61,62].
The assessment of the genetic structure of the giant kelp Macrocystis spp. across a broad latitudinal range in the HCS [55], reported low levels of genetic diversity in M. pyrifera populations and indicated the presence of a single species for this genus at a regional level. Later, this reduced genetic variation in M. pyrifera was reconfirmed from latitudes 12° S to 16° S. However, the presence of unique haplotypes was reported in populations from the San Lorenzo Island (Mpyr8), and from Los Bancos and San Nicolas Bay (Mpyr9) in Peru [61].
Regarding the Lessonia genus, the presence of two divergent lineages in central Chile was evidenced. For years, the existence of cryptic species was assumed; however, it was not after a few investigations [62,63,64], that the scientific name of L. berteroana started to be recognized for the northern populations, while L. spicata was kept mainly for southern ones. A recent molecular study showed that L. berteroana is distributed from at least 15°26′ S in Peru to 30° S in Chile [62].
2. Overall Search
The SALSA framework provided a large number of articles during the early stages of research. However, this was mainly completed according to the researchers’ lecture criteria when conducting the thematic analysis. Based on this, other methods, such as word-mining programs, are worth noting for future reviews.
Research on supporting services was the most prevalent (n = 59) and mostly related to ecological studies. This was followed by provisioning services (n = 19), most of them focused on fishery studies. Less attention was given to regulating (n = 3) and cultural services (n = 1). Only three papers discussed general topics related to all types of ES.
Publications predominantly addressed Chilean kelp forests (n = 77), with four publications explicitly mentioning ES in Chile [21,22,33,65]. Research on kelp forests in Peru (n = 5), or in Chile and Peru at the same time (n = 4), were extremely low. The number of publications showed a sustained increase during the last four decades, with more than half of the studies occurring in the last decade (2011–2022) (Figure 2).

Figure 2. Number of publications per decade.
3. General Ecosystem Services of Humboldtian Kelp Forests
In Central Chile, the study of ES using biomass, species richness and personal interviews [65,66] resulted in the identification of provisioning services (e.g., food), regulating services (e.g., biological production), supporting services (e.g., habitat or biodiversity) and cultural services. In northern Chile, an economic valuation of the ES provided by wild kelp populations of Lessonia spp. and M. pyrifera [22] indicated that kelp beds in this locality would have a value of USD 540 million per year over the next ten years with a constant annual increase. Of the total worth, 9% represented the service of the forests as an environmental buffer for CO2 capture or O2 production, 75% is provided by kelp fisheries and 15% by associated-species fisheries. The value of the total ES provided by the coastal benthic ecosystems of three bays (Mejillones, Antofagasta and Tongoy Bays) in northern Chile, including brown algae fisheries [21], was estimated to be about 8% of the total support value that ES provides to the regional economy. This shows the importance and role kelp forests have when providing numerous jobs, a source of income and food to coastal populations.
Kelp forests around the world support economic inputs, i.e., the value of kelp in South Africa is estimated at USD 434 million per year [67], and in the Falkland Islands at USD 342 billion per year [29]. The southeastern Pacific region’s kelp forests are dominated by M. pyrifera and their value in terms of ES has been evaluated at USD 811,000 per kilometer per year [22]. However, further research regarding the real extent of the species or assemblages are needed to generate accurate estimates in the HCS.
4. Supporting and Regulating Services
Kelp primary production enters the carbon cycle as wet biomass, detritus or dissolved organic matter, forming a food source for a wide range of organisms [17,68,69]. In general, ecosystems with a high net primary production generate more food, timber or fiber than less productive ecosystems [54,70]. As habitat formers, a single kelp sporophyte directly provides three distinct primary spaces: the holdfast, the stipe and the lamina. The morphological differences between Macrocystis pyrifera and Lessonia spp. kelp beds (i.e., stipe number, plant length, dichotomies per stipe and wet mass) influence the composition of the associated characteristic fauna and its functional relationships [69,71] (Figure 3). The kelp holdfast consists of a network of root-like ramifications, which provide galleries and crevices with a high structural complexity, allowing microenvironments to emerge as habitats for macroinvertebrate species such as echinoderms [45,72,73], crustaceans [74,75,76,77], polychaetes [76,78], bivalves [76], barnacles and limpets [45,79,80]. Fronds (blades and stipites) can form a dense canopy extending from the holdfast to the upper tip of the long stipes [78], representing habitats for various organisms, both epiphytes and associated fauna that seek shelter or food [36,81,82,83]. In kelp habitats, amphipods provide a link between kelp and higher trophic level species, including fish, which are voracious predators of amphipods [82,84]. Kelp sporophytes themselves are habitats for essential small-sized benthic suspension feeders which contribute to the recycling of nutrients (regulating services) [85]. Recently, a spatial optimization model to maximize the potential provision of ES was evaluated in coastal areas where Lessonia spp. was dominant, accounting for the role of dispersal and larval connectivity (regulating services). It was suggested that future modeling methodologies should encompass the diversity of coastal ecosystems and human activities to develop integrative spatial management [33].

Figure 3. Supporting services of kelp forests along the Humboldt Current System. The genus Lessonia usually forms a second canopy (a), while Macrocystis pyrifera forms an upper canopy (e). Kelp forests support habitats for a wide range of fauna, such as motile invertebrates (Pichidangui, Chile) (a); chondrichthyans that use kelp structures to deposit their capsules (Pucusana, Peru) (b); osteichthyes (Trachurus murphyi Nichols 1920; Pucusana, Peru) (c); and sessile organisms (Pucusana, Peru) (d,f).
A recent study integrating data of the macroinvertebrates associated with different forest forming species of Peru showed that more than 100 species are associated with M. pyrifera and L. trabeculata in central and southern Peru. Of these, L. trabeculata is the species with the highest diversity recorded [86]. Macroinvertebrate abundance, species richness and biomass significantly increased with holdfast size, explaining why Lessonia species have the highest associated diversity [45]. In Chile, at least 45 species were associated with Lessonia sp. [87,88] and 30 epifaunal invertebrate species inhabited M. pyrifera [80]. According to our search, L. trabeculata showed the highest number of taxa reported (n = 213) followed closely by M. pyrifera (n = 210). However, the number of phyla reported was higher in M. pyrifera (n = 17) than in Lessonia species (n = 7–13) (Figure 4). Overall, there were more reports of kelp-associated species for Chile than for Peru (Figure 5). The complete list of kelp-forest associated taxa reported per kelp species in Chile and Peru is shown in Table S2. The idea that this could be due to the high rates of speciation occurring in larger biogeographical provinces with lower surface temperatures and high endemism should be considered for further research [89,90].
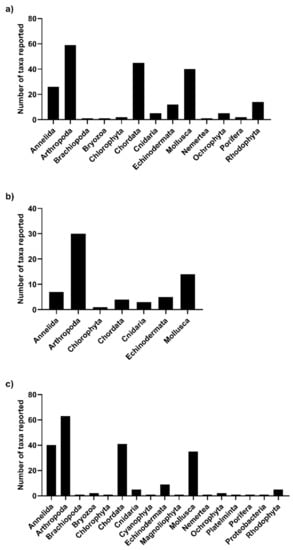
Figure 4. Number of kelp-forest associated taxa reported per Phyllum. (a) L. trabeculata; (b) L. berteroana/spicata; (c) M. pyrifera.

Figure 5. Number of kelp-forest associated taxa reported per kelp species in Chile (blue) and Peru (red).
The reports of fishes associated with kelp forests in southeastern Pacific are mainly from Chile [37,89,91,92,93,94,95,96,97,98,99,100] and show that kelp forests provide food and suitable habitats for benthic prey items through the understory community. It was suggested that understory habitats directly affect the diets of the fishes [93]. At least 25 species of reef fishes associated with Macrocystis pyrifera and Lessonia spp. were reported from the northern and central rocky coast of Chile, with many of them having socio-economic relevance at a local level [37,93]. Kelp forests provide food for many species. Experimental studies showed that the digestion of L. trabeculata is associated with the morphological features and the nutritional and reproductive status of the Zamba marblefish (Aplodactylus punctatus) [99].
Furthermore, kelp forests are strongly associated with food resources for coastal sharks, especially for males [101]. In contrast, pregnant females circle around vertical structures, selecting taller, physically stable and thicker sporophytes to anchor the tendrils of their capsules [39]. The redspotted catshark (Schroederichthys chilensis) has been associated with kelp forests dominated by L. trabeculata in Chile and Peru [39,101,102,103]. These consumers concurrently support even higher trophic level organisms, including predators such as seagulls [104], or the endangered sea otter Londra felina [92,105]. This is relevant for economies associated with the rich and productive HCS because previous studies have shown that biodiversity, including genetic diversity, is positively associated with the ES provided [106]. It is worth mentioning that our understanding of biodiversity may change over time, as new techniques are developed and integrated into ecological studies.
Regulating services have been mainly represented as the production of larvae that contributes to the regulation and stability of the marine ecosystem [106]. Multiple studies have highlighted the essential role of marine forests in larval dispersion and the colonization of distant habitats [57,73,106,107,108,109,110,111,112,113,114,115,116].
It is known that kelp detritus represents a subsidy of energy in low-productive habitats; hence, it is the main source of food for rich and abundant faunal assemblages, increasing the magnitude of carbon flow through consumers [60,75,106,107,108,109,110,111,112,113,114,115,116,117]. The latter coincides with experiments showing that trophic association with seaweeds is particularly important for epiphytic bryozoans under conditions of reduced particulate-food concentration [118]. Regarding kelp blue carbon studies, only one paper has been identified addressing the capacity of carbon storage by L. trabeculata in southern Peru [117]. Additionally, only one study addressed the economic value of the carbon that kelp assemblies capture in northern Chile [22].
5. Provisioning Services and Economical Benefits
In Chile and Peru, kelp species alone are a valuable bioresource used as raw material for alginate extraction [119]; feed for aquaculture species [120,121], and even stool pigeons [57]; organic fertilizer; biofuels; and human food [57,121,122,123] The use of kelp resources along the HCS is based on the harvest and collection of biomass, making Chile the leading producer country of raw material [120,121,122,123,124,125,126,127,128,129]. Kelp biomass is destined to alginate production, an industry valued at USD 213 million annually worldwide [129]. In northern Chile alone, more than 11,000 people depend directly or indirectly on the collection and harvesting of these resources [130]. For this reason, and to guarantee the sustainable production of kelps, alternative ways to manage and cultivate them are being investigated with the aim of obtaining these algae-associated benefits with lower ecosystem impacts [71,124,131].
Kelp fisheries are not the only kind of fisheries associated with kelp forests. Numerous fish, mollusks, crustaceans and other invertebrate species are associated with marine forests too, both in Chile and Peru [117,120,132]. Bioresources including Concholepas concholepas (Chilean abalone), Fisurella spp. (keyhole limpets), Loxechinus albus (red sea urchin), Pyura chilensis (red sea squirt), Octopus mimus (gould octopus) and various rock fishes, such as Cheilodactylus variegatus (Peruvian morwong), Paralabrax humeralis (Peruvian rock seabass), Pinguipes chilensis (Chilean sandperch) and Anisotremus scapularis (Peruvian grunt) have been reported in Humboldtian kelp forests. These species are continually captured by artisanal divers due to their socio-economic relevance, especially as food with high nutritional value [94,117,133,134].
6. Cultural Services
A collaborative paper between archaeologists and marine ecologists discussed the influence of kelp forests over the human migration from Asia to the Americas near the end of the Pleistocene. The study mentioned that marine forests provided protected nearshore areas for human migration, so it was easier for people to sail to the open sea. Kelp forests also provided food and materials that humans could keep for their sea voyages (e.g., kelp holdfasts were used for building boats) [135].
According to archaeological records, partially eaten and cooked seaweeds have been found at a 14,000-year-old site in Chile, suggesting that seaweed and associated fauna have been part of the human diet in the Western Hemisphere since ancient times [134,135,136,137]. In addition, remnants of algae, presumably Macrocrystis were found in tombs of the Nazca (10 BC–700 AD) and Paracas (700 BC–200 AD) cultures, revealing the preference for seaweeds in the diet and practices ancient coastal societies of Peru [58,138,139].
This entry is adapted from the peer-reviewed paper 10.3390/coasts2040013
This entry is offline, you can click here to edit this entry!