1. Epidemiological Studies
A number of epidemiological studies have shown that several dietary natural products and nutrients are associated with the risk of depression. For example, a prospective cohort study of 3177 Asian older adults showed that those who drank ≥3 cups of tea daily had a lower risk of depression (odds ratio (OR), 0.32; 95% confidence interval (CI), 0.12–0.84), compared with those who did not drink tea [
24]. Another study of 9576 Korean adults found ≥3 cups/week of green tea decreased the risk of self-reported lifetime depression [
25]. A cross-sectional survey found that a low intake of dietary fiber was associated with a high incidence of depression in hypertensive patients. Specifically, compared with those who consumed dietary fiber ≥ 15.4 g/day, the OR of 10.5–15.4 g/day to incidence of depression was 2.641 (95% CI, 1.050–6.640) [
26]. In addition, a cross-sectional study of 736 Brazilian farmers showed that the higher consumption of selenium decreased the risk of depression by 54% after adjusting for sociodemographic variables, pesticide intoxication, and lifestyle [
27]. A meta-analysis of 16 cross-sectional and 9 cohort studies showed that each 100 g increase of fruit intake decreased the risk of depression by 3% in cohort studies (risk ratio (RR), 0.97; 95% CI, 0.95–0.99), and each 100 g increase of vegetable consumption reduced the risk of depression by 5% in cross-sectional studies (RR, 0.95; 95% CI, 0.91–0.98) [
28]. Additionally, a higher intake of seaweed (OR, 0.38; 95% CI, 0.20–0.72) and mushroom fiber (OR, 0.18; 95% CI, 0.01–0.37) were negatively associated with depressive symptoms [
29]. Daily walnut consumption decreased the risk of depression with an OR of 0.67 (95% CI, 0.48–0.93), and this effect was found mainly in women (OR, 0.62; 95% CI, 0.46–0.84) but not in men (OR, 0.72; 95% CI, 0.41–1.27) [
30]. The exact reason for the gender difference remains unknown, and might have been a dose effect in which women consumed more walnuts. A cross-sectional study of 9183 Korean adults revealed that the prevalence of depression was decreased by 48% when people consumed ≥4 times of fish weekly, and the protective effect was particularly found in women (OR, 0.44; 95% CI, 0.29–0.67) but not in men (OR, 0.64; 95% CI, 0.30–1.37) [
31]. The reason for this gender difference might be biological differences between men and women, such as differences in sex hormones. Furthermore, a study showed there was a U-shaped relationship between fatty fish consumption and the risk of depression. In other words, moderate fatty fish consumption, but not high consumption, was associated with a lower risk of depression [
32]. Perhaps the high consumption of fatty fish may lead to obesity, which is associated with the occurrence of depression. Moreover, those who drank ≥4 cups of coffee daily had a lower risk of depression, compared with those who drank <1 cup/day (hazard ratios (HR), 0.37; 95% CI, 0.15–0.95) [
33]. A two-sample bidirectional Mendelian randomization analysis showed that there was a causal relationship between the higher consumption of carbohydrates and a lower risk of depression (OR, 0.42; 95% CI, 0.28–0.62) [
34].
It should also be pointed out that inconsistent results have been reported and not all studies showed protective effects of dietary natural products against depression. For example, a cross-sectional study found no link between legume and nut consumption and depression either in men (OR, 0.96; 95% CI, 0.54–1.71) or women (OR, 0.98; 95% CI, 0.65–1.48) [
35]. Soft drink is a kind of drink containing a great number of added sugars and mainly includes syrup, soda water, and other carbonated or non-carbonated beverages. A cross-sectional study of 8085 Chinese college students found that those who consumed >25 g sugar/day from soft drinks had an increased risk of depression compared with those who did not drink soft drinks [
36]. However, another study of 15,546 Spanish university graduates showed that there was no association between sugar-sweetened beverage consumption and the risk of depression (HR, 1.12; 95% CI, 0.90–1.41) [
37]. The inconsistent results from the different studies might be due to a racial factor (Chinese vs. Spanish).
In brief, many studies have pointed out that some dietary natural products could decrease the risk of depression, such as fish, walnuts, tea, and coffee. However, several studies have found no link between some dietary natural products and depression, such as legumes and nuts. In addition, inconsistent results regarding soft drink consumption and depression were reported by different studies, which should be further studied in the future. These results from epidemiological studies only indicated the possible anti-depressive effects of these substances, and clinical trials should be carried out to verify their effects on human beings. Furthermore, cell and animal experiments also should be performed to investigate the possible mechanisms of these substances on depression.
2. Experimental Studies
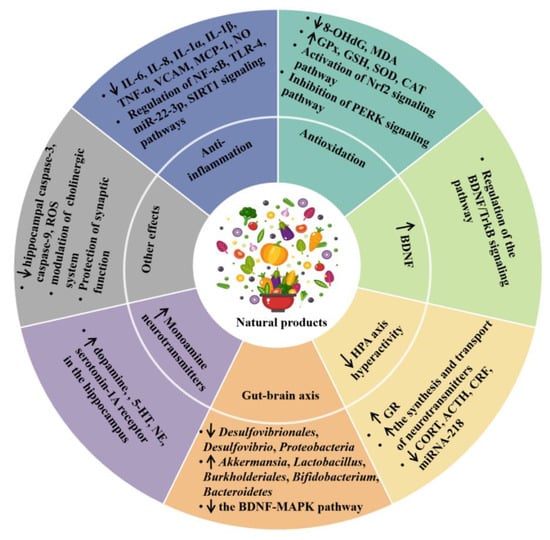
Many studies showed that some dietary natural products and nutrients had protective effects against depression through different mechanisms, including an anti-inflammatory effect, antioxidant effect, promoting the production of monoamine neurotransmitters, normalizing the hyperactivity of the HPA axis, regulating the microbiota–gut–brain axis, and other mechanisms [
7,
38,
39], which are discussed in detail below.
Figure 1. The effects and mechanisms of dietary natural products on depression. ↑ represents increase, and ↓ represents decrease. ACTH, adrenocorticotropic hormone; BDNF, brain-derived neurotrophic factor; CAT, catalase; CORT, corticosterone; CRF, corticotropin-releasing factor; GPx, glutathione peroxidase; GR, glucocorticoid receptor; GSH, glutathione; 5-HT, serotonin; IL, interleukin; MAPK, mitogen-activated protein kinase; MCP-1, macrophage chemoattractant protein 1; MDA, malondialdehyde; NE, norepinephrine; NF-κB, nuclear factor kappa-B; NO, nitric oxide; Nrf2, nuclear erythroid related factor 2; 8-OHdG, 8-hydroxydeoxyguanosine; PERK, protein kinase-like endoplasmic reticulum kinase; ROS, reactive oxygen species; SCFA, short-chain fatty acids; SIRT1, Sirtuin 1; TNF-α, tumor necrosis factor-α; SOD, superoxide dismutase; TrkB, tyrosine kinase receptor B; VCAM, vascular cell adhesion molecule.
2.1. Anti-Inflammatory Effect
Neuroinflammation is one of the most important risk factors for depression [
40]. Microglial cell activation could lead to neuroinflammation and increase the production of inflammatory cytokines, which results in neuronal damage and the progress of depression [
41]. It was found that n-3 polyunsaturated fatty acids (PUFA) could downregulate the expression of interleukin (IL)-6, IL-1β, tumor necrosis factor (TNF)-α, and prostaglandin E-2, which ameliorated post-menopausal depression induced by chronic mild stress and maternal separation [
42]. In addition,
Saccharina japonica is a common marine vegetable in East Asia and its ethanol extract decreased the depressive symptoms of dextran sodium sulfate-induced mice by increasing anti-inflammatory cytokine and downregulating expression levels of nuclear factor kappa-B (NF-κB), NOD-like receptor 3, and Toll-like receptor-4 (TLR-4) [
43]. Furthermore, apple phenolic extract improved lead acetate-induced depression-like behaviors through attenuating neuroinflammation and neuronal apoptosis, which was associated with the miR-22-3p/sirtuin 1 (SIRT1) signaling pathway [
39]. In an ovariectomy mice model, the oral application of
Prevotella histicola improved estrogen deficiency-induced depression through downregulating the expression levels of the vascular cell adhesion molecule (VCAM), macrophage chemoattractant protein 1 (MCP-1), IL-6, IL-8, and TNF-α in the ileum and colon of mice [
44]. Moreover,
Lacticaseibacillus paracasei NK112 exerted protective effects against
Escherichia coli-induced depression through decreasing the expression of IL-1α, IL-6, and TNF-α, and inhibiting the activity of NF-κB in the hippocampus [
45]. In a rat model of chronic unpredictable mild stress (CUMS)-induced depression, epigallocatechin gallate (EGCG) exerted an anti-depressive effect by reducing IL-6 and nitric oxide (NO) expression levels and decreasing the mRNA expression of caspase-3 and caspase-9 in the hippocampus [
46].
In a word, Saccharina japonica, apple phenolic extract, Prevotella histicola, Lacticaseibacillus paracasei NK112, n-3 PUFA, and EGCG could decrease depression through anti-inflammatory action in preclinical models, which should be verified in clinical trials.
2.2. Antioxidant Effect
Oxidative stress could disrupt the balance between oxidation and antioxidative defense and impair the structure and function of neural cells, which was closely associated with the development of depression [
47]. A number of studies have shown that some dietary natural products and nutrients have antioxidant activity that attenuates depression-like behaviors. For example, lycopene improved oxidative stress and endoplasmic reticulum stress-induced damage on neuroblastoma cells by reducing expression levels of 8-hydroxydeoxyguanosine, malondialdehyde (MDA), and protein carbonyls and inhibiting the protein kinase-like endoplasmic reticulum kinase (PERK) signaling pathway [
48].
Grewia asiatica berry is a popular berry in Pakistan that is mainly consumed in the form of carbonated drinks and fresh juices and is rich in phenols, anthocyanins, vitamin C, and flavonoids [
49].
Grewia asiatica berry juice could improve depressive symptoms by decreasing oxidative damage in the brain through increasing superoxide dismutase (SOD) and glutathione peroxidase (GPx) levels [
50]. Another study pointed out that
Saccharina japonica ethanol extract decreased the depression-like behaviors of dextran sodium sulfate-induced mice by increasing the activity of SOD [
43]. In addition, maqui berries had an anti-depressive effect against post-stroke depression by upregulating expression levels of reduced glutathione (GSH) and enhancing the activities of SOD and catalase (CAT) [
51]. Moreover, fish oil was useful in the prevention of depression in old MRL/
lpr mice through enhancing nuclear erythroid related factor 2 (Nrf2)-mediated antioxidant defense [
52].
In brief, Grewia asiatica berry juice, Saccharina japonica, maqui berry, fish oil, and lycopene mitigated depression by antioxidant action in preclinical models, which should be further studied on human beings.
2.3. Modulating the Production of Monoamine Neurotransmitters
The monoamine hypothesis is one of the most generally accepted etiological hypotheses of depression, and modulating monoamine neurotransmitter systems plays an important role in the treatment of depression [
53]. Substantial studies have shown that some dietary natural products and nutrients could regulate the production of monoamine neurotransmitters to decrease depression. For instance, deoiled sunflower seeds ameliorated the depression-like behaviors of CUMS-induced mice through upregulating levels of dopamine, serotonin, and norepinephrine [
54]. Moreover, another study showed that EGCG improved depression-like behaviors induced by CUMS through reducing the level of serotonin in the colon but increasing it in the hippocampus [
55]. Furthermore, a study showed navel orange essential oil exerted anti-depressive effects via increasing serotonin and dopamine levels in the brain [
56]. In addition, adzuki bean sprout fermented milk could decrease the symptoms of mild depression by upregulating the expression levels of serotonin, norepinephrine, and dopamine in the hippocampus [
57]. Additionally, n-3 PUFA had preventive effects against depression by increasing brainstem serotonin levels and the hippocampal expression of the serotonin-1A receptor [
42].
In a word, deoiled sunflower seeds, navel orange essential oil, adzuki bean sprout fermented milk, EGCG, and n-3 PUFA had preventive and therapeutic effects on depression via the modulation of monoamine neurotransmitter production, and clinical trials should be carried out to verify the potential effects and mechanisms of these substances on human beings.
2.4. Promoting the Production of Neurotrophins
Neurotrophins are a family of functionally and structurally related proteins, which play a crucial role in promoting the survival, development, and function of neurons, including BDNF, nerve growth factor, neurotrophin-3, and neurotrophin-4 [
58]. Several studies have shown that some dietary natural products could increase the production of neurotrophins to improve depression. For instance, garlic essential oil showed an anti-depressive effect against CUMS-induced depression via upregulating the expression levels of hippocampal BDNF, cyclic adenosine monophosphate response element-binding protein (CREB), and protein kinase B [
59]. Moreover,
Geum japonicum is a popular medicinal herb in Asia. It showed neuroprotective effects on corticosterone (CORT)-induced depressive mice by upregulating the expression of BDNF in the hippocampus. It also decreased CORT-induced neurotoxicity in SH-SY5Y cells [
60]. In a lupus-prone MRL/
lpr old mice model of depression, fish oil and conjugated linoleic acid all increased the expression levels of BDNF and synaptic protein in the brain [
52]. Additionally, purified anthocyanin from purple cauliflower improved the depressive symptoms of CUMS-induced mice by suppressing the activity of monoamine oxidases and upregulating expression levels of monoamine neurotransmitters, BDNF, and tyrosine kinase receptor B (TrkB) [
19]. In a mouse model of CUMS-induced depression, sesamin improved depressive symptoms by increasing neurotrophin expression levels, such as BDNF and neurotrophin-3 [
61]. Furthermore, a study found that supplements with probiotics ameliorated the depression-like behaviors of rats with epilepsy through upregulating the expression levels of BDNF and nerve growth factor [
62]. In a rat model of male hypogonadism, resveratrol alleviated depression-like behaviors via increasing hippocampal and prefrontal cortical levels of BDNF and neurotrophin-3 [
63].
In short, Geum japonicum, garlic essential oil, fish oil, conjugated linoleic acid, and anthocyanin could ameliorate depression through increasing the production of BDNF, nerve growth factor, and neurotrophin-3, and their effects and mechanisms should be further investigated by clinical trials.
2.5. Inhibition of the HPA Axis Hyperactivity
The HPA axis is an important part of the neuroendocrine system and plays an essential role in the control of stress response and the mediation of mood [
64]. The hyperactivity of the HPA axis inhibited the negative feedback signal of cortisol and upregulated the levels of corticotropin-releasing factor (CRF) and adrenocorticotropic hormone (ACTH), which further resulted in depressive symptoms [
38]. It was found that eicosapentaenoic acid (EPA)-enriched phospholipids suppressed HPA axis hyperactivity to ameliorate depressive symptoms induced by chronic stress and lipopolysaccharide (LPS) [
65]. Additionally, saponin compounds extracted from the traditional Chinese medicine Baihe Zhimu Tang showed an anti-depressive effect through inhibiting the hyperactivation of the HPA axis and improving the synthesis and transport processes of neurotransmitters [
66]. In a rat model of post-menopausal depression induced by chronic mild stress and maternal separation, n-3 PUFA exerted antidepressant-like effects through reducing blood levels of CORT and ACTH, decreasing the brain expression of CRF and miRNA-218, and increasing the expression of the glucocorticoid receptor [
42]. Furthermore, a study showed n-3 PUFA improved pup separation-induced postpartum depression via regulating the HPA axis by reducing circulating levels of ACTH and CORT and downregulating the expression of hypothalamic CRF [
67]. In addition, royal jelly (a common dietary supplement) attenuated CUMS-induced depression by inhibiting the biosynthesis of CORT in the adrenal gland [
68].
In brief, royal jelly, EPA-enriched phospholipids, saponin compounds, and n-3 PUFA reduced depressive symptoms by suppressing the hyperactivity of the HPA axis in animal models, which should be certified in clinical trials.
2.6. Modulation of Microbiota–Gut–Brain Axis
Nowadays, the important role of gut microbiota in the prevention and management of various diseases has increasingly been recognized [
69,
70,
71,
72,
73,
74,
75]. Many studies have shown that disorders of gut microbiota affected the production of cytokines and other inflammatory factors, which regulated several signaling pathways associated with depression [
76]. In addition, gut microbiota disorders increased gut barrier permeability and stimulated immune responses and systemic inflammation, which affected the activity and function of the HPA axis and the efficacy of monoamine neurotransmitters [
10]. A number of studies have pointed out that several probiotics could improve the symptoms of depression. For example,
Prevotella histicola was an emerging probiotic and showed a protective effect against estrogen deficiency-induced depression by elevating the abundance of intestinal flora, especially
Akkermansia and
Lactobacillus [
44]. Moreover,
Lactobacillus casei exerted anti-depressive effects on postpartum depression via changing the composition of gut microbiota, increasing the expression level of BDNF, and suppressing the activity of the BDNF–MAPK pathway [
77]. In addition, a study showed that
Lactobacillus casei mitigated CUMS-induced depressive symptoms through reversing the structure change of gut microbiota and regulating BDNF/TrkB signaling pathways [
78]. Another study showed that
Bifidobacterium E41 and M2CF22M7 suppressed depression-like behaviors via improving the microbial dysbiosis and enhancing expression levels of serotonin and BDNF [
79]. Moreover,
Lactobacillus kefiranofaciens ZW3 isolated from Tibetan Kefir grains could improve the symptoms of depression through regulating the gut microbiota composition and ameliorating constipation via increasing the content of fecal water [
80]. In addition,
Lactobacillus gasseri NK109 improved
Escherichia coli K1-induced depression through decreasing the expression level of IL-1α in activated macrophages and regulating gut microbiota via vagus nerve-mediated gut–brain signaling [
81].
In addition to the probiotics mentioned above, it was found that other dietary natural products and nutrients could also regulate the gut–brain axis to decrease depression-like behaviors. For example, the high consumption of dietary fiber ameliorated antenatal obesity-induced postpartum depressive-like behaviors, and the mechanisms were related to increasing the expression levels of norepinephrine and serotonin, inhibiting neuroinflammation, promoting the formation of short-chain fatty acids, and the reconstruction of the gut microbiome [
82]. In a rat model of ACTH-induced depression, chlorogenic acid exerted anti-depressive effects by increasing the relative abundance of
Burkholderiales and
Bifidobacterium and reducing the relative abundance of
Desulfovibrionales and
Desulfovibrio [
83]. Furthermore,
Bifidobacteria-fermented red ginseng exerted protective effects against
Escherichia coli-induced depression by enhancing the abundance of
Bacteroidetes, decreasing the abundance of
Proteobacteria, and upregulating the expression of BDNF mediated by NF-κB [
84]. In addition, the disorder of gut microbiota was correlated with circadian rhythm disorders, and tea polyphenols were found to regulate the circadian rhythm and increase the abundance of probiotics to attenuate depressive symptoms [
10]. Nicotinamide riboside is a form of vitamin B
3 and is mainly found in milk and yeast [
85]. In a mouse model of alcohol-induced depression, nicotinamide riboside changed the composition of gut microbiota, which further decreased the expression levels of inflammation-related cytokines and increased BDNF levels in the hippocampus [
86]. Additionally, soy isoflavones improved the depressive symptoms of CUMS rats, and the mechanisms were associated with enhancing the diversity of gut microbiota and upregulating the monoamine neurotransmitters levels [
87]. Coniferyl ferulate is a phenolic acid compound mainly found in umbelliferous plants and exerted a protective effect against CUMS-induced depression by improving the reconstruction of the gut microbiome and downregulating the expression levels of IL-6, IL-1β, and TNF-α to decrease colonic inflammation [
88].
Semen sojae praeparatum is a traditional fermented food and showed anti-depressive effects through modulating the microbiota–gut–brain axis. Specifically,
Semen sojae praeparatum upregulated the abundance of the genus
Ruminococcaceae_UCG-008 and regulated serotonin, norepinephrine, GABA, and BDNF content in the hippocampus [
89].
In a word, some dietary natural products and nutrients could attenuate depression through regulating the microbiota–gut–brain axis in preclinical models, such as Lactobacillus casei, Bifidobacterium E41 and M2CF22M7, Lactobacillus kefiranofaciens ZW3, Lactobacillus gasseri NK109, Prevotella histicola, Semen sojae praeparatum, Bifidobacteria-fermented red ginseng, dietary fiber, tea polyphenols, chlorogenic acid, nicotinamide riboside, soy isoflavones, and coniferyl ferulate. The possible effects and mechanisms of these substances on depression should be further studied in clinical trials.
2.7. Other Mechanisms
In addition to the related mechanisms mentioned above, several dietary natural products and nutrients also exerted an anti-depressive effect through other mechanisms. For instance,
Grewia asiatica berry juice reduced the depression-like behaviors of rats through the modulation of the cholinergic system via decreasing levels of acetylcholinesterase and MDA [
50]. In a mouse model of subchronic and mild social defeat stress-induced depression, heat-killed
Lactobacillus helveticus strain MCC1848 ameliorated depressive symptoms by improving the alteration of gene expression in nervous system development and signal transduction [
90]. Furthermore, lotus plumule is the green embryo of lotus seeds and is widely used for tea in China. The alkaloids extracted from lotus plumule mitigated LPS-induced depressive symptoms through suppressing BDNF-mediated endoplasmic reticulum stress and increasing autophagy [
91]. In addition, a study found that curcumin attenuated the depression-like behaviors of LPS-induced rats through suppressing excessive synaptic loss and improving synaptic function [
92]. Additionally, short-chain fatty acids are the gut microbial metabolites and have neuroprotective bioactivity [
93]. A study found that short-chain fatty acids decreased depressive-like behaviors of high fructose-fed mice via inhibiting microglia activation and reducing blood–brain barrier damage [
94].
In short, Grewia asiatica berry juice, heat-killed Lactobacillus helveticus strain MCC1848, alkaloids from lotus plumule, and curcumin could ameliorate depression, and the potential mechanisms included the modulation of the cholinergic system, suppression of mitochondrial apoptotic pathways, inhibition of BDNF-mediated endoplasmic reticulum stress, improvement of synaptic function, and promotion of nervous system development and signal transduction. In the future, these mechanisms should be widely studied, and also should be verified based on human beings.
3. Clinical Trials
A placebo-controlled and randomized study of 25 healthy college students showed that daily drinking apple cider vinegar could reduce the risk of depression, which might be related to inhibit activation of hexosamine pathway and promote metabolism of glycine, serine and threonine [
97]. Another double-blind, randomized and controlled clinical trial of 60 women with depressive disorder found that supplement with flaxseed oil twice a day for 10 weeks increased the concentration of serum BDNF and attenuated depressive symptoms [
98]. In addition, a randomized controlled pilot study of 143 postpartum women showed that frequently drinking magnolia tea had protective effects against postpartum depression [
99]. In a randomized double-blind and placebo-controlled trial of 64 healthy adolescents, it was found that daily supplement with wild blueberry had positive effects on the prevention of depression [
100]. Furthermore, a randomized, double-blind, placebo-controlled trial of 82 patients with depression showed that the consumption of
Lacticaseibacillus paracasei strain Shirota for 9 weeks improved depression-like behaviors and ameliorated constipation of patients, which was related to upregulate the beneficial
Adlercreutzia,
Megasphaera and
Veillonella levels [
101]. In a randomized trial of 24 healthy volunteers, consumption of yogurt twice daily after lunch and dinner did not significantly change biomarker levels of depression, but when the participants regularly consumed yogurt and daily exercised, the level of serotonin increased [
102]. Additionally, a meta-analysis of 36 clinical trials including 2788 participants showed that the higher flavonoid consumption improved depressive symptoms (Mean difference = −1.65; 95% CI, −2.54, −0.77) [
103]. Moreover, a meta-analysis of 23 randomized controlled trials including 237 participants revealed the saffron improved the symptoms of depression, compared with placebo [
104].
In brief, clinical trials have found that saffron, wild blueberry, flaxseed oil, magnolia tea, apple cider vinegar, Lacticaseibacillus paracasei strain Shirota and flavonoid could decrease depression. Although these results from clinical trials are more reliable than those from preclinical studies, more randomized, double-blind, placebo-controlled trials should be carried out for verifying their effects on different race persons in different places of world.
This entry is adapted from the peer-reviewed paper 10.3390/antiox11112132