Enhancement of terrestrial carbon (C) sequestration on marginal lands in Canada using bioenergy crops was investigated. This study quantified the long-term C sequestration potentials at the system-level in nine-year-old (2009–2018) woody (poplar clone 2293–29 (Populus spp.), hybrid willow clone SX-67 (Salix miyabeana)), and herbaceous (miscanthus (Miscanthus giganteus var. Nagara), switchgrass (Panicum virgatum)) bioenergy crop production systems on marginal lands in Southern Ontario, Canada.
Woody crops and switchgrass were able to increase SOC significantly over the tested period. However, when long term soil organic carbon (∆SOC) gains were compared, woody and herbaceous biomass crops gained 11.0 and 9.8 Mg C ha−1, respectively, which were not statistically different. Results also indicate a significantly higher total C pool [aboveground + belowground + soil organic carbon] in the willow (103 Mg ha−1) biomass system compared to other bioenergy crops. In the nine-year study period, woody crops had only 1.35 Mg C ha−1 more system-level C gain (SLCG), suggesting that the influence of woody and herbaceous biomass crops on SLCG and ∆SOC sequestrations were similar. Further, among all tested biomass crops, willow had the highest annual SLCG of 1.66 Mg C ha−1 y−1.
- root biomass
- system-level C gain
- orthogonal contrast
- carbon stock
- coil health
- climate change mitigation
1. Carbon Stock Allocation in Plant Components
Results of the analysis of variance (ANOVA) for stem and leaf, aboveground and belowground (roots) C stock of biomass in 2017 and 2018 for different bioenergy cropping systems in southern Ontario, Canada are presented in Table 1. Quantified aboveground biomass C stock ranged from 4.01 Mg C ha−1 in poplar to 7.41 Mg C ha−1 in willow (Cycle 3, Year 2), and in switchgrass, it was 4.44 Mg C ha−1 in 2017 (Table 2). In 2018, aboveground biomass C stock ranged from 4.38 Mg C ha−1 in switchgrass to 12.39 Mg C ha−1 in willow (Cycle 3, Year 3) (Table 2). Belowground biomass C stock also ranged from 4.11 Mg C ha−1 in poplar to 10.06 Mg C ha−1 in willow (Cycle 3, Year 3) (Table 2).
Table 1. Significance level (p-value) for the analysis of variance (ANOVA). Source of variations include plant and block as main effects on stem and leaf carbon stock (stem, leaf), aboveground (AGB) and belowground (BGB) carbon stock in biomass in 2017 and 2018 in biomass cropping systems, southern Ontario, Canada.
| Source of Variation | df | Stem 2017 2 | Leaf 2017 2 | AGB 2017 2 | Stem 2018 3 | Leaf 2018 | AGB 2018 | BGB 2018 |
|---|---|---|---|---|---|---|---|---|
| Plant 1 | 3 | 0.0103 | 0.0068 | 0.0079 | 0.0038 | <0.0001 | 0.0021 | 0.0013 |
| Woody vs. Herbaceous | 1 | 0.4114 | <0.0001 | 0.1147 | 0.1835 | <0.0001 | 0.0462 | 0.5174 |
| Woody (W. vs. P.) | 1 | 0.0157 | 0.0380 | 0.0176 | 0.0195 | <0.0001 | 0.0080 | 0.0002 |
| Herbaceous (M. vs. S.) | 1 | NA | NA | NA | 0.0025 | 1.0000 | 0.0037 | 0.4674 |
| Block CV (%) R-Square | 5 | 0.1400 | 0.3776 | 0.1499 | 0.3533 | 0.5841 | 0.3485 | 0.2521 |
| 23.27 | 53.30 | 25.00 | 28.20 | 40.83 | 27.84 | 24.65 | ||
| 0.81 | 0.81 | 0.82 | 0.73 | 0.92 | 0.76 | 0.79 |
Table 2. Aboveground (2017 and 2018) and belowground (2018) biomass carbon stock in bioenergy crops and averages for woody and herbaceous cropping systems on marginal lands in southern Ontario, Canada. Standard errors are in brackets.
| Cropping System | Aboveground Biomass (Mg C ha−1) 2017 1 | Aboveground Biomass (Mg C ha−1) 2018 2 | Belowground Biomass (Roots) (Mg C ha−1) 2018 | ||||
|---|---|---|---|---|---|---|---|
| Stems 3 | Leaves | Total | Stems 3 | Leaves | Total | ||
| Poplar | 3.62 (0.43) b | 0.45 (0.04) b | 4.01 (0.42) b | 7.01 (0.98) b | 0.57 (0.11) b | 7.65 (1.08) bc | 4.11 (0.45) b |
| Willow | 6.37 (0.79) a | 1.10 (0.21) a | 7.41 (1.00) a | 10.83 (0.79) a | 1.54 (0.14) a | 12.39 (0.91) a | 10.06 (0.81) a |
| Switchgrass | 4.44 (0.27) b | 0.00 (0.00) b | 4.44 (0.27) b | 4.38 (0.60) b | 0.00 (0.00) c | 4.38 (0.60) c | 8.45 (1.34) a |
| Miscanthus | n/a | n/a | n/a | 10.91 (1.94) a | 0.00 (0.00) c | 10.91 (1.94) ab | 7.45 (1.16) a |
| Woody | 5.00 (0.60) x | 0.77 (0.14) x | 5.74 (0.72) x | 8.92 (0.82) x | 1.06 (0.18) x | 10.02 (0.98) x | 7.09 (1.00) x |
| Herbaceous | 4.44 (0.27) x | 0.00 (0.00) y | 4.44 (0.27) x | 7.65 (1.55) x | 0.00 (0.00) y | 7.65 (1.55) y | 7.95 (0.84) x |
C stock allocation in different plant components among all four biomass crops is presented in Figure 1. Results indicate that for poplar, 60.15% of total tree C was stored in stems and branches, and 4.88% in leaves, while 34.97% was stored in the roots. In poplar, leaves, stems, and branches together stored in total 65.03% of the plant C, which is the highest proportion of C within aboveground biomass quantified among all tested woody and herbaceous biomass crops in this study. For willow, 48.25% of the total tree C was stored in stems and branches, and 6.95% in leaves, while 44.80% of C was allocated to belowground roots. In this context, switchgrass and miscanthus C allocations to belowground components were 65.86% and 40.6% C, respectively, while 34.2% and 59.4% of the total C was stored in the aboveground components.
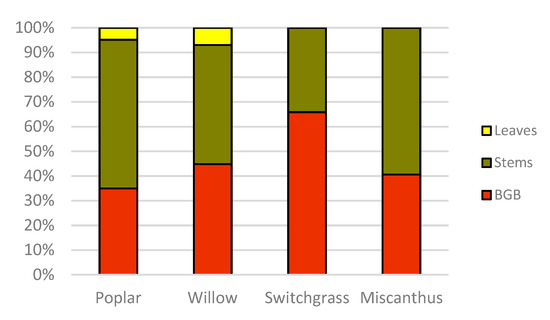
Figure 1. Carbon stock allocation to plant components (leaves litter in three-year-old woody crops, stems and leaves in herbaceous, stems and branches in woody crops and belowground biomass) in nine-year-old (2018) bioenergy cropping systems in southern Ontario, Canada. In herbaceous crops “stems” included both leaves and stem carbon stock.
2. Soil and Total Carbon Pools in Woody and Herbaceous Bioenergy Cropping Systems
Results indicate that woody biomass systems (willow and poplar) and switchgrass significantly increased SOC between 2009 (baseline) to 2018 (Table 3), while miscanthus failed to reach significance. In this context, the 2018 SOC sequestration values were not significantly different across all biomass cropping systems (Table 4). The average SOC in 2018 in woody and herbaceous cropping systems were 80.1 and 73.8 Mg ha−1, respectively (Table 5). SOC sequestration gains (∆SOC) for switchgrass, willow, poplar, and miscanthus were 11.8, 11.7, 10.3, and 7.7, Mg ha−1, respectively (Table 5).
Table 3. Significance level (p-value) from the t-test (α = 0.05) on soil total organic carbon sequestration or accumulation since 2009 (baseline) to 2018 in different biomass cropping systems on marginal lands in southern Ontario, Canada.
| Cropping System | P (T <= t) Two-Tail |
|---|---|
| Poplar | 0.040 |
| Willow | 0.050 |
| Switchgrass | 0.034 |
| Miscanthus | 0.143 |
Table 4. Significance level (p-value) for the analysis of variance (ANOVA). Source of variations include plant and block as main effects on total organic carbon sequestration (SOC) in 2009 and 2018 and long-term soil carbon sequestration between 2009–2018 (∆SOC), total carbon pool at the system-level in 2018 (CPool) and system-level carbon gain (SLCG) in biomass cropping systems, southern Ontario, Canada.
| Source of Variation | df | SOC 2009 | SOC 2018 | ∆SOC | CPool | SLCG |
|---|---|---|---|---|---|---|
| Plant 1 | 3 | 0.7623 | 0.2165 | 0.6682 | 0.0130 | 0.5062 |
| Woody vs. Herbaceous | 1 | 0.3046 | 0.0619 | 0.3831 | 0.0200 | 0.3341 |
| Woody (W. vs. P.) | 1 | 0.9166 | 0.7939 | 0.7401 | 0.0098 | 0.4273 |
| Herbaceous (M. vs. S.) | 1 | 1.0000 | 0.3818 | 0.4331 | 0.7628 | 0.3959 |
| Block | 5 | 0.5440 | 0.0497 | 0.1921 | 0.0447 | 0.1758 |
| CV (%) | - | 9.24 | 8.20 | 67.74 | 6.92 | 55.13 |
| R-Square | - | 0.40 | 0.66 | 0.47 | 0.75 | 0.50 |
Table 5. Soil organic carbon (SOC) measurements (2009 and 2018) in 0–30 cm depth, long-term soil organic carbon sequestration between 2009–2018 (∆SOC), total carbon pool at the system-level in 2018 (CPool) in bioenergy crops and averages for woody and herbaceous cropping systems on marginal lands in southern Ontario, Canada. Standard errors are in brackets.
| Cropping System | SOC (2009) (Mg C ha−1) | SOC (2018) (Mg C ha−1) | ∆SOC (Mg C ha−1) | CPool 1 (Mg C ha−1) |
|---|---|---|---|---|
| Poplar | 69.3 (2.08) a | 79.6 (3.66) a | 10.3 (4.30) a | 91.4 (4.11) b |
| Willow | 68.9 (2.69) a | 80.6 (4.32) a | 11.7 (3.11) a | 103.1 (3.49) a |
| Switchgrass | 64.0 (3.17) a | 75.9 (0.39) a | 11.8 (3.46) a | 88.7 (1.00) b |
| Miscanthus | 64.0 (3.17) a | 71.8 (3.32) a | 7.7 (1.59) a | 90.2 (4.91) b |
| Woody Crops | 69.1 (1.62) x | 80.1 (2.70) x | 11.0 (2.54) x | 97.2 (3.12) x |
| Herbaceous Crops | 64.0 (2.07) x | 73.8 (1.73) x | 9.8 (1.92) x | 89.4 (2.33) y |
In this study, the total C pool, at the system-level, was calculated by combining all C pools for woody and herbaceous crop (SOC + above- and belowground C stock) in year of 2018. Summing all these C pools, yielded a total C pool of 91.4, 103.1, 88.7, and 90.2 Mg ha−1 for poplar, willow, switchgrass, and miscanthus, respectively (Table 5). The total C pool was significantly different between woody and herbaceous systems (Table 4). The average of the total C pools was 97.2 and 89.4 Mg ha−1 in woody and herbaceous cropping systems, respectively (Table 5).
3. System-Level Carbon Gain in Woody and Herbaceous Bioenergy Cropping Systems
Quantified SLCGs from 2009 to 2018 were compared among all four bioenergy cropping systems (Table 4, Figure 2) and ranged from 9.7 Mg ha−1 for miscanthus to 14.9 Mg ha−1 for willow. The average SLCG in woody and herbaceous cropping systems from 2009 to 2018 was 13.3 and 11.8 Mg ha−1, respectively (Figure 2), which were not significantly different (Table 4). In the same period, when we consider biomass crop types, switchgrass and willow gained 4.4 and 3.3 Mg ha−1 more system-level carbon in their respective systems compared to miscanthus and poplar, respectively (Figure 2).
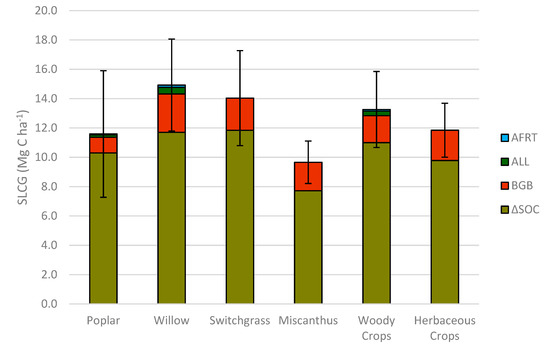
Figure 2. System-level carbon gain in bioenergy crops in southern Ontario, Canada, during 2009 to 2018, as well as averages for woody and herbaceous cropping systems. System-level carbon gain delineated by long-term soil organic carbon from 2009 to 2018 (∆SOC), belowground biomass carbon (BGB), annual leaf litter (ALL) input carbon, and annual fine root turn-over (AFRT) carbon.
4. Discussion
4.1. Carbon Stock Allocation in Plant Components
Data from this study show that among the woody species, willow aboveground biomass C stock was significantly (p < 0.05) higher than poplar, and among the herbaceous species, miscanthus aboveground biomass C was significantly (p < 0.05) higher than switchgrass (mean difference 4.7 Mg C ha−1 between willow and poplar and 6.5 Mg C ha−1 between miscanthus and switchgrass). However, belowground biomass C stock was not significantly different between woody and herbaceous crops (Table 1 and Table 2). The annual C accumulation found in our woody cropping systems (3.34 and 0.79 Mg C ha−1 yr−1 biomass C accumulation for above- and belowground biomass, respectively) were consistent with previously reported values by Oliveira et al. [1] that demonstrated accumulation of carbon in the belowground fraction of the biomass in poplar short rotation plantations under Mediterranean conditions ranged from 0.86 to 0.91 Mg C ha−1 yr−1, whereas the aboveground carbon accumulation ranged from 3.89 to 6.48 Mg C ha−1 yr−1. It should be explained that, in our study, the woody biomass was harvested every three years (three-year cycle) and therefore, the total aboveground biomass carbon was divided by three to calculate annual accumulation and the belowground biomass carbon was divided by nine (total age of the stand, between 2009 and 2018, Table 2). However, the stand maturity in Oliveira et al. [1] study was only three to four years. Another study by Verlinden et al. [2] recorded aboveground C accumulation as 2.5 Mg C ha−1 yr−1 under low productivity conditions. Other studies have also reported similar values (e.g., 3.1–5.75 Mg C ha−1 yr−1 [3]).
In the literature, for short rotation willow and poplar plantations, belowground biomass accumulation has been reported as 1.8–3.5 Mg ha−1 y−1 for first rotation (e.g., [4]) and 2.4 Mg ha−1 y−1 for a five-year-old (second cycle) short rotation [5]. Similar to the results derived from this study (Table 2), Coleman et al. [6] have also reported that the mean belowground biomass for willow and poplar clones were 16.51 and 8.79 Mg ha−1 (7.88 and 4.19 Mg C ha−1 y−1), respectively. Zan et al. [7] have reported an annual belowground C in the root biomass for switchgrass and willow in southwestern Quebec, Canada, over a three-year period to be 1.06 and 1.25 Mg C ha−1 y−1, respectively. These numbers are comparable to the numbers derived in this study (0.94 and 1.12 Mg C ha−1 y−1 for switchgrass and willow, respectively, Table 2). However, their numbers may be slightly higher than those reported in our study because they measured belowground biomass up to 60 cm soil depth compared to the 30 cm soil depth in our study. Therefore, results from perennial biomass studies suggest that if unproductive agricultural lands in Canada are converted to perennial biomass crops, irrespective of the type of biomass crop (herbaceous or woody), considerable amounts of atmospheric C can be stored in belowground components of the crop.
Results from this study suggest that the higher allocation of C in leaves in willow provides a potential for increased C inputs to soil via litterfall contributing to soil organic carbon (SOC) sequestration. Our findings also indicate that herbaceous biomass crops can contribute to SOC sequestration enhancement. Collectively, belowground biomass C allocations quantified in all biomass crops tested in this study suggest that they can contribute significantly to enhance SOC sequestration over the years, as they are perennial in nature. Agostini et al. [8] have reported that annual C inputs from miscanthus roots were about half that of switchgrass, given similar turnover time. However, in our study (Table 2), belowground biomass C was not significantly different between miscanthus and switchgrass, with values of 8.45 and 7.45 Mg C ha−1 for switchgrass and miscanthus, respectively. It is also worth noting that roots of herbaceous crops contributed, on an average, more than half of the total plant C (53.2%) compared to woody crop roots (39.9% of total tree C). This is particularly important as the C stock of roots have been neglected and not been estimated in most studies ([9][10] from [11]).
4.2. Soil and Total Carbon Pools in Woody and Herbaceous Bioenergy Cropping Systems
Both woody biomass cropping systems and the switchgrass system were able to increase SOC significantly (p < 0.05) in the nine-year period, while miscanthus failed to reach significance. However, when 2018 SOC sequestration values were tested across all biomass cropping systems, results indicate that SOC values were not significantly different from each other (Table 4), while willow sequestrated numerically the highest amount of SOC at 80.6 Mg ha−1 (Table 5).
It is interesting to note that the difference in SOC sequestration (∆SOC) between the woody and the herbaceous systems was only 1.2 Mg ha−1 between 2009 and 2018, which was not significant. Therefore, results from this study suggest that ∆SOC in perennial biomass systems may not be influenced by the biomass type.
In Coleman et al. [6], the conventional agricultural field having corn-bean-wheat crop rotation in GARS, Guelph, Ontario, Canada, recorded a SOC stock of 63.80 ± 7.04 Mg ha−1 at 0 to 30 cm soil depth. This agricultural field is adjacent to the biomass research plots of our study having the same soil type, (Figure 1). Comparing the 2009 baseline SOC stock value for the bioenergy cropping systems to that of the adjacent agricultural field SOC stock value (Coleman et al. [6]) implies that in the agricultural field, SOC stock recorded in 2016 is close to our baseline SOC stock value measured in 2009. Therefore, any SOC stock gain between 2009 and 2018 in our biomass study is likely to be attributed to the presence of perennial bioenergy cropping system. Based on the above SOC numbers (Coleman et al. [6]) it is assumed that SOC in the previous management system will not have changed unless a sustainable management strategy such as biomass crops are implemented.
Root C input from belowground biomass and fine-root turnover can significantly contribute to increases in SOC under woody and switchgrass cropping systems [7][12]. Liebig et al. [13] monitored switchgrass bioenergy production in central and northern Great Plains, USA for a five-year period and reported that the cropping system significantly affected change in SOC and increased it at a depth of 0–30 cm at a rate of 1.1 Mg C ha−1 year−1 (4.0 Mg CO2 ha−1 year−1). In contrast to switchgrass [7], miscanthus has root crowns, resulting in a reduction in the proportion of fine roots compared to switchgrass, which could contribute to reduced SOC gains over the years. This could have been the reason as to why SOC gain was low for miscanthus compared to switchgrass in this study (Table 5).
Based on total C pool values, it is obvious that the willow system has the largest C pool compared to all tested biomass systems (Table 5). The total C pool in woody crop systems was also significantly higher (97.2 Mg ha−1) than herbaceous cropping systems (89.4 Mg ha−1). This finding is supported by other studies which report that the annual net SOC storage change exceeds the minimum mitigation requirement under perennial energy crops by far [8]. Similarly, Carvalho et al. [14] have suggested that if the current bioethanol sector (such as corn and sugarcane) is changed to bioenergy feedstocks with more allocation to belowground C it could increase soil C stocks at a much faster rate. Other reports have also shown that the proportion of the total system C (biomass + root + SOC) in a willow bioenergy system was 14.4% and 15.6% more than in switchgrass and corn systems, respectively [15].
It is interesting to note that in Table 5, total C pool was significantly (p < 0.05) higher in the willow system compared to all other tested biomass crops in this study. This is mainly due to significantly (p < 0.05) higher aboveground biomass C (Table 2) and numerically higher belowground biomass C (Table 2) compared to other tested biomass crops. Willow has the ability to coppice more vigorously after each harvest than poplar and also adapts itself better than other biomass crops on low-productive or marginal lands [16]. The total C pool values reported in this study are within the previously reported range of values, 12–175 Mg C ha−1 [11][17][18].
4.3. System-Level Carbon Gain in Woody and Herbaceous Bioenergy Cropping Systems
Differences in SLCGs were not significantly affected by bioenergy crops (Table 4). However, considering the numerical values, results suggest that when we compare SOC pools of all tested woody and herbaceous biomass systems, willow and switchgrass are sequestering C in the soil in measurable quantities (Table 5, Figure 2).
These results indicate that willow cropping systems are able to gain 1.66 Mg ha−1 carbon annually at the system-level, which is the highest C gain among all other bioenergy crops in this study on marginal lands in southern Ontario, Canada. This can be attributed to the higher contribution of coarse and fine root C, as well as annual leaf litter in willow system. However, slowly decomposable rhizomes are incorporated into soil organic matter in herbaceous cropping systems. Carvalho et al. [14] concluded that the belowground biomass C pool plays a critical role in building and maintaining SOC, especially due to rhizodeposition inputs and the higher potential of C retention rates. Chemical composition of belowground biomass can also explain the higher C retention rate as roots have a higher concentration of phenolic and lignaceous compounds [19], and enhance soil aggregation, which increases the physical protection of organic C added into the soil [20].
Coleman et al. [6] reported an increase of 1.16 Mg C ha−1 y−1 for SOC stocks in concentrated short-rotation woody production systems (willow and poplar), which is supporting the results derived in this study. Several studies [7][8][21] show that deciduous woody cropping systems contribute greater levels of SOC than herbaceous systems due to leaf litter inputs and higher rate of root turnover. The fine roots in willow cropping systems are often a focal point of belowground C sequestration because they represent a vital biomass C of the total willow root C [22]. Soil depth and texture should be considered when comparing soil C inputs from belowground biomass in woody and herbaceous bioenergy crops. This study was conducted in a marginal sandy-loam textured soil with a depth of only 0 to 30 cm. However, soil C sequestration up to a depth of 1 m will provide more value, especially when dealing with perennial biomass cropping systems [23]. Therefore, SOC data from this study are likely underestimating the total soil C pool. In a 30-year simulation study the belowground inputs to soil C from miscanthus were 34% of the total inputs, and it could be as high as 60% when considering a rooting depth of 1 m [14].
Empirical and modeled studies also suggest that increases in soil clay content can reduce root contributions to soil C and in such cases, it is more likely that aboveground C inputs dominate the top 30 cm of soil C over time [14][22][24]. The 30-year simulation study in a soil with 29% clay showed a net increase in the soil C pool of 5.7 Mg ha−1, where root systems were responsible for 46% of the total inputs. However, in a very clayey soil, root systems had a lower total contribution to the soil C pool (18%) and the main portion of this input resulted from mortality and incorporation of roots and rhizomes during soil preparation at replanting [14].
5. Conclusions
Overall, results from this study suggest that both woody biomass cropping systems (poplar and willow) and switchgrass are able to increase SOC significantly in the nine-year period of this study (2009–2018). However, carbon gains at the system-level (SOC + Root C + Leaf litter C + Fine Root C), did not significantly differ between woody and herbaceous systems during the study period (Figure 2). The findings show that the woody systems may have an advantage over herbaceous biomass systems based on the numerical SLCG values. It will be interesting to monitor as to how these systems will differ in their C sequestration numbers into the future as both, woody and herbaceous systems, are considered to be productive for up to 22 years [16].
In terms of aboveground biomass carbon, the assessed woody systems are producing significantly higher aboveground biomass carbon than herbaceous crops. However, we did not include this in the system-level carbon sequestration calculation as the aboveground biomass will be utilized for bioenergy production, bio-products, or for other purposes such as animal bedding, garden mulch, etc. In contrast, this study has demonstrated that the amount of belowground biomass C (roots) in herbaceous or in woody biomass crops was similar in both systems, with a difference of only 0.9 Mg ha−1. However, at the ‘system-level’, we have taken into account carbon gains associated with above- and belowground biomass such as, leaf-litter inputs, fine-root turnover, and belowground biomass carbon additions in roots. If carbon credits are calculated for biomass crops, biomass removal from the system and taking it out of the ‘farm gate’ for energy production will not be considered as system-level gain. However, any other carbon additions to the system that are related to aboveground biomass (for example leaf-litter), as indicated above, should be accounted for and we have done so in this study.
Above- and belowground biomass C and SOC contributed to the total C pool, which was significantly higher in woody bioenergy cropping systems and highest overall in the willow system. Therefore, these findings demonstrate that the willow bioenergy cropping system sequestered significantly more atmospheric CO2 than all other tested systems, suggesting it has the potential for producing bioenergy with the lowest net CO2 emissions.
Across Canada, it is estimated that there are approximately 9.5 million ha of potentially useable marginal lands, and there are close to 1 million ha in Ontario alone [25]. Given the willow cropping systems’ C gain per year (1.66 Mg C ha−1 y−1), if this system is established on all 9.5 million ha of marginal non-agricultural lands in Canada, the maximum per year gain will be more than 15,770 Gg C y−1 or 57,876 Gg CO2 y−1, which is 8% of the total annual Canadian GHGs emissions (716,000 Gg CO2 y−1) [26]. Therefore, based on our results, there is considerable potential for terrestrial C sequestration in Canada and Ontario by converting low quality agricultural lands to biomass production.
This entry is adapted from the peer-reviewed paper 10.3390/su12093901
References
- N. Oliveira; R. Rodríguez-Soalleiro; César Pérez-Cruzado; I. Cañellas; H. Sixto; R. Ceulemans; Above- and below-ground carbon accumulation and biomass allocation in poplar short rotation plantations under Mediterranean conditions. Forest Ecology and Management 2018, 428, 57-65, 10.1016/j.foreco.2018.06.031.
- Melanie Verlinden; L.S. Broeckx; Donatella Zona; Gonzalo Berhongaray; Toon De Groote; Marta Camino-Serrano; Ivan A. Janssens; Reinhart Ceulemans; Net ecosystem production and carbon balance of an SRC poplar plantation during its first rotation. Biomass and Bioenergy 2013, 56, 412-422, 10.1016/j.biombioe.2013.05.033.
- Shengzuo Fang; J. Xue; L. Tang; Biomass production and carbon sequestration potential in poplar plantations with different management patterns. Journal of Environmental Management 2007, 85, 672-679, 10.1016/j.jenvman.2006.09.014.
- R. D. Hangs; J. J. Schoenau; K. C. J. Van Rees; N. Belanger; Timothy A. Volk; Leaf Litter Decomposition and Nutrient-Release Characteristics of Several Willow Varieties Within Short-Rotation Coppice Plantations in Saskatchewan, Canada. BioEnergy Research 2014, 7, 1074-1090, 10.1007/s12155-014-9431-y.
- Renato S. Pacaldo; Timothy A. Volk; Russell D. Briggs; Greenhouse Gas Potentials of Shrub Willow Biomass Crops Based on Below- and Aboveground Biomass Inventory Along a 19-Year Chronosequence. BioEnergy Research 2013, 6, 252-262, 10.1007/s12155-012-9250-y.
- Coleman, B.; Bruce, K.; Chang, Q.; Frey, L.; Guo, S.; Tarannum, M.S.; Bazrgar, A.; Sidders, D.; Keddy, T.; Gordon, A.; et al. Quantifying C stocks in high-yield, short-rotation woody crop production systems for forest and bioenergy values and CO2 emission reduction. For. Chron. 2018, 94, 260–268.
- Claudia S Zan; James W. Fyles; Patrick Girouard; Roger A Samson; Carbon sequestration in perennial bioenergy, annual corn and uncultivated systems in southern Quebec. Agriculture, Ecosystems & Environment 2001, 86, 135-144, 10.1016/s0167-8809(00)00273-5.
- Agostini, F.; Gregory, A.S.; Richter, G.M. Carbon Sequestration by Perennial Energy Crops: Is the Jury Still Out? Bioenergy Res. 2015, 8, 1057–1080.
- Joel N. Swisher; Cost and performance of CO2 storage in forestry projects. Biomass and Bioenergy 1991, 1, 317-328, 10.1016/0961-9534(91)90012-2.
- Jon Unruh; Ra Houghton; Pa Lefebvre; Carbon storage in agroforestry: an estimate for sub-Saharan Africa. Climate Research 1993, 3, 39-52, 10.3354/cr003039.
- Peichl, M.; Thevathasan, N.V.; Gordon, A.M.; Huss, J.; Abohassan, R.A; Carbon sequestration potentials in temperate tree-based intercropping systems, southern Ontario, Canada. Agrofor. Syst. 2006, 66, 631-643, .
- A B Frank; J. D. Berdahl; J. D. Hanson; M.A. Liebig; H. A. Johnson; Biomass and Carbon Partitioning in Switchgrass. Crop Science 2004, 44, 1391-1396, 10.2135/cropsci2004.1391.
- Mark A. Liebig; M. R. Schmer; K. P. Vogel; R. B. Mitchell; Soil Carbon Storage by Switchgrass Grown for Bioenergy. BioEnergy Research 2008, 1, 215-222, 10.1007/s12155-008-9019-5.
- Joao L. N. Carvalho; Tara Hudiburg; Henrique C. J. Franco; Evan H. DeLucia; Contribution of above- and belowground bioenergy crop residues to soil carbon. GCB Bioenergy 2017, 9, 1333-1343, 10.1111/gcbb.12411.
- D. Andress; Bioenergy crops and carbon sequestration. CRC. Crit. Rev. Plant Sci. 2005, 24, 1–21, .
- Floriane Marsal; Naresh Thevathasan; Solène Guillot; John Mann; Andrew M. Gordon; Mahendra Thimmanagari; William Deen; Salim Silim; Raju Soolanayakanahally; Derek Sidders; et al. Biomass yield assessment of five potential energy crops grown in southern Ontario, Canada. Agroforestry Systems 2016, 90, 773-783, 10.1007/s10457-016-9893-3.
- Dixon, R.K. Agroforestry systems: Sources of sinks of greenhouse gases? Agrofor. Syst. 1995, 31, 99–116.
- Schroeder, P; Carbon storage benefits of agroforestry systems. Agrofor. Syst. 1994, 27, 89–97, .
- Bolinder, M.A.; Angers, D.A.; Giroux, M.; Laverdière, M.R. Estimating C inputs retained as soil organic matter from corn (Zea Mays L.). Plant Soil 1999, 215, 85–91.
- Oades, J.M. An overview of processes affecting the cycling of organic carbon in soils. In The Role of Non-Living Organic Matter in the Earth’s Carbon Cycle; Zepp, R.G., Sonntag, C.H., Eds.; Wiley: New York, NY, USA, 1995; pp. 293–303.
- Bransby, D.I.; McLaughlin, S.B.; Parrish, D.J; A review of carbon and nitrogen balances in switchgrass grown for energy. Biomass Bioenergy 1998, 14, 379–384, .
- Beyhan Amichev; Ryan D. Hangs; Sheala M. Konecsni; Christine N. Stadnyk; Timothy A. Volk; Nicolas Bélanger; Vladimir Vujanovic; Jeff Schoenau; Judicaël Moukoumi; Ken C.J. Van Rees; et al. Willow Short-Rotation Production Systems in Canada and Northern United States: A Review. Soil Science Society of America Journal 2014, 78, S168-S182, 10.2136/sssaj2013.08.0368nafsc.
- Kristina J. Anderson-Teixeira; Michael D. Masters; Christopher Black; Marcelo Zeri; Mir Zaman Hussain; C. Bernacchi; Evan H. DeLucia; Altered Belowground Carbon Cycling Following Land-Use Change to Perennial Bioenergy Crops. Ecosystems 2013, 16, 508-520, 10.1007/s10021-012-9628-x.
- Osaki, M.; Shinano, T.; Matsumoto, M.; Ushiki, J.; Shinano, M.M.; Urayama, M.; Tadano, T; Productivity of high-yielding crops: V. Root growth and specific absorption rate of nitrogen. Soil Sci. Plant Nutr. 1995, 41, 635–647, .
- Tingting Liu; Ted Huffman; Suren Kulshreshtha; Brian McConkey; Yuneng Du; Melodie Green; Jiangui Liu; Jiali Shang; Xiaoyuan Geng; Bioenergy production on marginal land in Canada: Potential, economic feasibility, and greenhouse gas emissions impacts. Applied Energy 2017, 205, 477-485, 10.1016/j.apenergy.2017.07.126.
- Environment and Climate Change Canada. Canadian Environmental Sustainability Indicators: Progress towards Canada’s Greenhouse Gas Emissions Reduction Target. 2020. Available online: https://www.canada.ca/en/environment-climate-change/services/environmental-indicators/progress-towards-canada-greenhouse-gas-emissions-reduction-target.html (accessed on 9 May 2020).