1. Introduction
The plant kingdom is the richest source of naturally occurring phenolic compounds, as there are about 8000 phenolic compounds in its distribution [
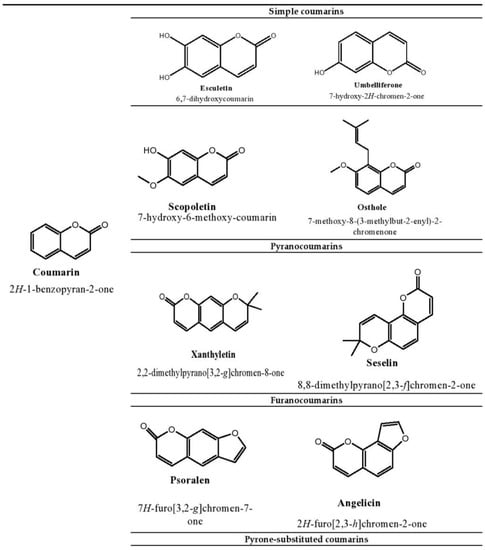
1]. The presence of these phenolic compounds in nature has been extensively used by researchers to tackle disease stratification. Coumarins are one of the main bioactive phenolic or heterocyclic compounds with their large distribution in nature. These compounds belong to the benzopyrone class and are found in various plants, notably in the seeds of the tonka bean and
Dipteryx odorota. These phenolic compounds are classified into four types, namely, simple coumarins, furanocoumarins, pyranocoumarins, and pyrone-substituted coumarin [
2]. Umbelliferone, esculetin, scopoletin, and osthole are some examples of simple coumarin compounds. Coumarin derivatives that are hydroxylated and alkoxylated are classified as simple coumarins. The origin of furanocoumarins involves the association of the five-membered heterocyclic furan ring with coumarins, while their analogs are called pyranocoumarins, which are six-membered heterocyclic non-aromatic compounds containing five carbon atoms. Psoralen, angelicin, and xanthyletin are some examples of furanocoumarins, while seselin is a member of pyranocoumarins. Warfarin is a potent anti-coagulant derivative of coumarin and is classified in pyrone-substituted coumarins [
2] (
Figure 1).
Figure 1. Structure of coumarin compounds.
Several reports have highlighted that some anticancer drugs used in the treatment of tumorigenesis not only kill malignant cells but also affect normal cells by producing reactive oxygen species that subsequently damage deoxyribonucleic acid (DNA), proteins, and lipids [
3,
4]. For example, doxorubicin is a potent anti-cancer drug that causes apoptosis in malignant cells by accumulating hydroxyl radicals, resulting in the development of oxidative stress in the body [
5]. 2-Methoxyestradiol is another drug that exhibits anti-cancer properties by activating the c-Jun-N-terminal kinase (JNK) signaling pathway-induced apoptosis [
6,
7]. In addition, this drug has been found to induce hydrogen peroxide (H
2O
2) in mitochondria [
8]. An in vitro study by Heo et al. reported that resveratrol induces cell-cycle arrest in melanoma cells (A375SM cells) by activating the reactive oxygen species (ROS)-mediated P53–P38 and ER stress pathway [
9]. These studies have shown that drugs used in cancer therapy can act as a source of oxidative stress in the body.
Inflammation recruits the immune cells by liberating the chemokines and cytokines toward the site of oxidative stress [
10]. Increased production of ROS by polymorphonuclear neutrophils at the site of inflammation leads to tissue injury and endothelial dysfunction [
11]. Likewise, an increase in ROS and reactive nitrogen species increased the level of isoprostanes and prostaglandins (PGE), which in turn caused rheumatoid arthritis (RA) [
12]. Published reports claim that patients with asthmatic symptoms have higher levels of ROS [
13,
14]. This demonstrated that the generation of free radicals can lead to degenerative diseases such as asthma.
The etiology of arthritis Is not completely understood [
15]; however, it is implied that activation of nuclear factor-kappa B (NF-κB) and mitogen-activated protein kinase (MAPK) signaling pathways are major contributors to this disease. Phosphorylated IkB can cause NF-κB translocation to the nucleus in response to lipopolysaccharides (LPS) stimulation, where it binds to target genes and produces inflammatory cytokines [
16,
17]. Similarly, the release of pro-inflammatory cytokines such as tumor necrosis factor-α (TNF-α), interleukin (IL)-1, and matrix metalloproteinase (MMP) enzymes such as MMP-2, -3, and -9 cause cartilage and joint deterioration in arthritis [
18,
19]. Conversely, oxidative stress is thought to be a primary contributor to the persistent inflammation prevalent in arthritis cartilage. In osteoarthritis (OA), the level of inflammatory mediators such as IL-1β, TNF-α and IL-6 are increased, which directs the induction of reactive oxygen species and causes matrix breakdown and joint deformation. Since ROS and inflammation are interconnected, they become excellent therapeutic targets [
20].
Besides cancer and inflammation, diabetic patients are found to have a low level of antioxidant enzymes that trigger a cascade of hyperglycemia-induced oxidative stress [
21,
22]. Diabetic nephropathy is one of the serious complications associated with diabetic individuals. Together, oxidative stress and inflammation alter lipid and protein functions and induce glycoxidation in diabetes [
23,
24], suggesting the role of ROS in diabetes and its associated complications.
Fat accumulation has been reported in individuals with non-alcoholic fatty liver conditions, and those with obesity, high blood sugar, and cholesterol are at great risk of developing such conditions [
25,
26,
27,
28,
29,
30,
31,
32]. In the liver, ROS such as superoxide anion radicals and H
2O
2 are present which promotes the pathogenesis of oxidative stress [
33]. A high level of ROS leads to changes in the structure of proteins, lipids, and DNA, which consequently accumulates the ruptured macromolecules and causes liver injury such as fatty liver [
34]. Conversely, high levels of by-products of lipid peroxidation (hydroxyoctadecadienoic acid and hydroxyeicosatetraenoic acid) are elevated in the liver, with an increase in triglyceride, suggesting the role of free radicals and high-fat diet (HFD) in the development of fatty liver by inducing the lipid peroxidation [
35].
Carbon tetrachloride is a common agent for lipid peroxidation, which can cause liver damage [
36]. Cytochrome P450 2E1 is an enzyme that enables the conversion of carbon tetrachloride to trichloromethyl radicals to initiate lipid peroxidation in the body and signals the lipids to produce oxidation products that ultimately hinder the functioning of the liver by developing a condition called liver injury [
37].
These facts showed that apoptosis, imbalance of glycemic index, hepatic failure, and the generation of free radicals are highly correlated with each other. It is therefore imperative to interrupt the link between cancer, inflammatory disorders, arthritis, diabetes, and hepatic diseases from oxidative stress to prevent the stratification of diseases.
Esculetin (6,7-dihydroxychromen-2-one) is a coumarin derivative that structurally contains the two hydroxyl groups at the 6th and 7th carbon atoms. The presence of more hydroxyl groups in esculetin helps scientists to replace hydroxyl groups with any group to prepare a new derivative against disease outbreaks. The bioactivities and therapeutic applications of coumarin compounds and their derivatives depend on their structural arrangement. Free radicals act as a potent source for the pathogenesis of many diseases, whereas the presence of hydroxyl groups on esculetin makes this compound more efficient to act as an antioxidant by inhibiting the oxidative stress in disease conditions [
38,
39,
40]. It has been found that the attachment of the hydroxyl group to phenolic compounds can effectively connect with free radicals [
41]. These hydroxyl groups in phenolic compounds can also exhibit chelation with transient metals such as copper and iron [
42]. Esculetin exhibits dual modulation of apoptosis, as well as anti-diabetic and anti-inflammatory action that may be partly attributed to its antioxidant characteristic.
2. Therapeutic Applications of Esculetin
Oxidative stress plays an important role in the pathogenesis of chronic ailments such as cancer, diabetes, liver disease, and inflammatory disorders. Esculetin exhibits numerous therapeutic applications such as free radical scavenging, suppression of dysregulated transcription factors in cancer, inhibition of inflammatory pathways involved in arthritis, management of glycemic index, fatty liver disease, etc.
2.1. The Role of Esculetin in Cancer Treatment
Cancer is a lethal condition that arises as a result of abnormal cell proliferation in the body and leads to organ failure and death. Many developments have been made either to control or cure this lethal disease. Coumarin compounds have always been considered the first choice for researchers because of their excellent biological activity and low toxicity [
45]. A study by Arora et al. showed that esculetin at 100 µM arrests the growth of cancer cells in the G1 phase of the cell cycle. It impedes the binding interaction between nuclear factor erythroid 2-related factor 2(Nrf2) and Klech-like-ECH-Associated Protein-1 (KEAP-1) by activating the antioxidant response element (ARE) pathway and attenuating the NF-κB activity leading to apoptosis in the human pancreatic cancer cell (PANC-1) [
46]. Esculetin at 20 µM inhibits leukemia cell proliferation and induces autophagy through the formation of autophagic vesicles. In addition, it downregulates the expression of cyclin D1, D3, DK4, and DK2, causing a cell-cycle arrest at the G0/G1 phase. This coumarin compound was shown to block the phosphorylation of MAPK and extracellular signal-regulated kinase (ERK), thereby inhibiting the activation of Raf/MAPK/ERK signaling [
47], and was also reported to downregulate the JNK and ERK pathways in leukemia U937 cells at 30 µg/mL, suggesting its potential in inhibiting the tumorigenesis [
48].
Benzo[a]pyrene is known to play an important role in lung cancer [
49], and 50 mg/kg esculetin treatment showed an arrest in the cancerous cell proliferation by downregulating Bcl-2 and NF-κB, causing apoptosis [
50]. Esculetin at 55 µg/mL concentration not only inhibits pancreatic and lung cancer but also prevents colon cancer by activating MAPK signaling pathways, caspase-3 and 9, leading to apoptosis. It was also found to release cytochrome c into the cytosol by increasing the depolarization of the mitochondrial membrane and increasing the B-cell lymphoma-2-associated x protein expressions [
51]. In the oral squamous cancer cell, 20 µg/mL of esculetin has been shown to downregulate the expression of specificity protein 1 (Sp1), p27, cyclin D1, Mcl-1, and survivin, thus inducing apoptosis [
52]. In larynx cancer, 2 and 10 µM esculetin treatment was found to inhibit the Janus kinase–signal transducer and activator of transcription (JAK/STAT) pathway by suppressing the phosphorylation of STAT3 and subsequent translocation into the nucleus. Furthermore, esculetin causes cell-cycle arrest at the G1/S phase and thus supports apoptosis [
53].
In hepatocellular carcinoma, esculetin at 2.24 µM promotes apoptosis by arresting the cells at the S-phase of the cell cycle. In addition, esculetin was found to elevate the mechanism of caspase-3 and 9 and reduced the mitochondrial membrane potential. Esculetin significantly increased the Bax expression, thereby reducing the Bcl-2 expression, thus exhibiting its anti-cancerous potential [
54]. Overactivation of insulin-like growth factor-1/phosphoinositide-3-kinase/Protein kinase B (IGF1/PI3K/Akt) and IGF1/MAPK signaling pathways contributed to the development of tumors; however, esculetin at 850 µM was found to diminish the mitochondrial membrane potential while simultaneously activating the mitochondrial apoptotic pathway in the MGC-803 gastric cancer cell line. It increased the cytochrome c release from mitochondria, Bax/Bcl-2 index, and activated the activity of caspase-3 and 9 by suppressing the IGF-1/PI3K/Akt pathway [
55]. Esculetin at 200 µg/mL was reported to prevent the proliferation, migration, and invasion of renal carcinoma cells by cell-cycle arrest at G0/G1 and G2/M phase, downregulating the expression of Cyclin D1, CDK4, CDK6, and cellular myelocytomatosis (c-Myc) resulting in apoptosis. Levels of E-cadherin were increased, whereas the expressions of N-Cadherin and vimentin were downregulated with esculetin treatment [
56] (
Table 1,
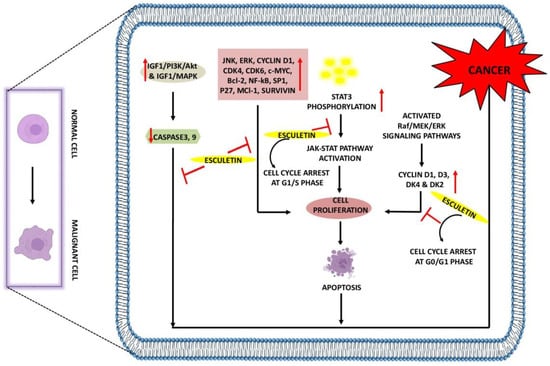
Figure 2).
Figure 2. Anti-cancer effects of esculetin: Esculetin downregulates the IGF1/PI3K/AKT, IGF1/MAPK, JNK, ERK, NF-κB, P27, Survivin, and Bcl-2. Esculetin also inhibits cell proliferation by inhibiting the STAT3 phosphorylation and cyclin D1, D3, DK4, and DK2, leading to apoptosis and suppression of cancer. ↑: Overexpression; ↓: low expression; ⊣: inhibition.
Table 1. Mechanism and function of esculetin in treatment of cancer.
| Pharmacological Mechanism |
Inhibition/Activation/
Downregulation/Upregulation |
Model Used |
Dosage |
Application |
Reference |
| |
Cell-cycle arrest at G1-phase
Activate ARE pathway and impede binding interactions between Nrf2 and KEAP-1
Attenuate NF-κB pathway |
Human PANC-1 cells |
100 µM |
In vitro |
[45] |
| |
Inhibit cell proliferation
Induce autophagy by forming
autophagic-vesicles
Downregulate cyclin D1, D3,
DK4 and DK2
Induce cell-cycle arrest at G0/G1-phase
Block MEK/ERK phosphorylation by inhibiting Raf/MEK/ERK signaling |
Human leukemia cells (HL-60 cells) |
20 µM |
In vitro |
[47] |
| |
Downregulate JNK/ERK signaling |
Human leukemia cells (U937 cells) |
30 µM |
In vitro |
[48] |
| |
Downregulate Bcl-2 and NF-κB
expressions
Induce apoptosis |
Benzo[a]pyrene-induced lung carcinogenesis in Swiss-albino mice |
50 mg/kg |
In vivo |
[50] |
| Anti-cancer |
Activate MAPK signaling
Activate caspase-3 and 9 and cause
apoptosis
Release cytochrome c into cytosol
Increase mitochondrial membrane
depolarization
Increase Bax expression |
Human colon cancer cells (HT-29 cells) |
55 µg/mL |
In vitro |
[51] |
| |
Suppress SP1, p27, cyclin D1, Mcl-1, survivin expressions
Induce apoptosis |
Oral squamous cancer (HN22 and HSC4 cells) |
20 µg/mL |
In vitro |
[52] |
| |
Downregulate STAT3 phosphorylation
Inhibition of JAK/STAT pathways
Induce cell-cycle arrest at G1/S-phase |
Laryngeal cancer (Hep2 cells) |
2, 10 µM |
In vitro, In vivo |
[53] |
| |
Cell-cycle arrest at S-phase
Elevate caspase-3, 9 expressions
Reduce mitochondrial membrane
potential
Increase Bax expression
Downregulate Bcl-2 expression |
Hepatocellular carcinoma (C57BL/6 mice were implanted with Hepa1–6 cells and SMMC-7721 cells) |
2.24 mM |
In vitro, In vivo |
[54] |
| |
Suppress IGF-1/PI3K/Akt and IGF-1/MAPK signaling
Reduce mitochondrial membrane
potential
Release cytochrome c from
mitochondria
Increase Bax, Bcl-2, caspase-3, 9
Expressions |
Human gastric cancer (MGC-803 and GES-1 cells) |
850 µM |
In vitro |
[55] |
| |
Inhibit proliferation, migration and
invasion of renal cancerous cells
Induce cell-cycle arrest at G0/G1 and G2-phase
Downregulate cyclin D1, CDK4, CDK6 and c-Myc expressions
Increase E-cadherin level by decreasing N-cadherin and vimentin expressions |
Renal carcinoma (786-O and SN12-PM6 cells) |
200 µg/mL |
In vitro |
[56] |
2.2. The Role of Esculetin in Oxidative Stress Treatment
Free radicals are the single unpaired electron species that exist independently in the body and are capable of causing oxidative damage to DNA, proteins, and carbohydrates. Therefore, antioxidants act as an eminent tool against oxidative damage. Many coumarin compounds, including esculetin, have been evaluated using in vitro and in vivo models of oxidative stress. H
2O
2 is known to play a crucial role in generating oxidative stress. Treatment of 5 µM esculetin has been reported to increase the phosphorylation ofNrf2 and the expression of NAD(P)H:quinone oxidoreductase1(NQO1) in C2C12 myoblasts cells. In addition, esculetin at the same dose was also reported to activate the ERK signaling pathway and exhibit protective effects against H
2O
2-induced free oxidative stress [
57]. Likewise, Kim and colleagues showed that Chinese hamster lung fibroblast cells (V79-4 cells) treated with H
2O
2 showed an increase in lipid peroxidation free radical generation, leading to DNA damage, whereas the free radical scavenging and intracellular ROS scavenging activity of esculetin were found to be 77% and 75% at 10 µg/mL [
58]. Increased serum levels of alkaline phosphatase (ALP), AST, and ALT in carbon tetrachloride-induced injury in rat liver were reduced with 35 mg/kg esculetin dosage, which was mainly due to its lipid peroxidation and free radical inhibition activity [
59].
In Alzheimer’s disease, 20 µM esculetin activates the Nrf2 and increases the glutathione levels in SH-SY5Y cells. In addition, esculetin increases the phosphorylation of ERK and Akt and thus protects cells from oxidative stress-induced damage by amyloid proteins [
60]. In human dermal fibroblast cells, esculetin at 0.6 and 2.1 µg/mL concentrations effectively scavenges the superoxide and DPPH radicals and inhibits MMP-1 [
61]. It has been found that H
2O
2 upregulates the expression of MMP-1, thus promoting skin aging and oxidative stress by activating the MAPK and AP1 signaling pathways. The treatment of 5 µg/mL esculetin to H
2O
2-induced oxidative stress in HaCaT cells results in the inhibition of MMP-1, phospho-MEK1, phospho-ERK1/2, phospho-SEK1, and phospho-JNK1/2 along with the intracellular Ca
2+ levels [
62]. In addition, esculetin at 50 µL was reported to protect human fibroblast from oxidative stress-induced DNA damage induced by linoleic acid hydroxide and iron (III) ion [
63] (
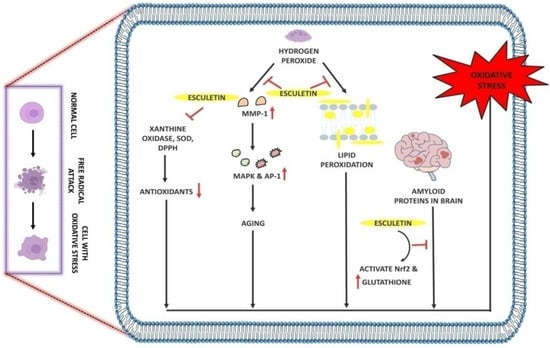
Table 2,
Figure 3).
Figure 3. Antioxidant effect of esculetin: Esculetin blocks the activity of H2O2 which further inhibits the MMP-1 and lipid peroxidation in cell membranes. Esculetin prevents the brain from amyloid-induced oxidative stress via activating the Nrf2 and increasing the levels of glutathione, thus inhibiting oxidative stress. ↑: Overexpression; ↓: low expression; ⊣: inhibition.
Table 2. Mechanism and function of esculetin in treatment of oxidative stress.
| Pharmacological Mechanism |
Inhibition/Activation/
Downregulation/Upregulation |
Model Used |
Dosage |
Application |
Reference |
| Antioxidant |
Increase phosphorylation of
Nrf2 and NQO1
Activate ERK signaling pathways
Show protective effect against H2O2-induced oxidative stress |
H2O2-induced oxidative stress in C2C12 myoblasts cells |
5 µM |
In vitro |
[57] |
Scavenge DPPH, hydroxyl and
intracellular ROS
Inhibit lipid peroxidation,
protein carbonyl and DNA-damage induced by H2O2 |
Chinese hamster lung fibroblast cells (V79-4 cells) |
10 µg/mL |
In vitro |
[58] |
Scavenge free radicals
Inhibition of lipid peroxidation, AST, ALT and ALP in liver |
CCl4-induced acute hepatotoxicity in male Sprague Dawley rats |
35 mg/kg |
In vivo |
[59] |
Activate Nrf2
Increase phosphorylation of ERK
signaling and Akt signaling
Increase glutathione levels |
Amyloid protein-induced oxidative stress and neuronal death in SH-SY5Y cells |
20 µM |
In vitro |
[60] |
Inhibit DPPH, Xanthine oxidase,
superoxide radicals
Downregulate MMP-1 expression |
Oxidative stress in human dermal fibroblasts cells (HDF-cells) |
0.6 µg/mL and
2.1 µg/mL |
In vitro |
[61] |
Inhibit phospho-MEK1,
phospho-ERK1/2, phospho-SEK1 and phospho-JNK1/2 along with
intracellular Ca2+ levels
Inhibit MMP-1 expression |
H2O2-induced oxidative stress in Human HaCaT keratinocytes cells |
5 µg/mL |
In vitro |
[62] |
| |
Scavenge hydroxyl radicals and protect DNA from oxidative damage |
Lipid-hydroperoxide-induced oxidative damage in human diploid fibroblast cells (TIG-7 cells) |
50 µL |
In vitro |
[63] |
2.3. The Role of Esculetin in Inflammation Treatment
Inflammation is defined as the defense mechanism that occurs in response to infection or injury and helps the body to maintain homeostasis. During infection or injury, the damaged cell triggers the release of various physiological messengers including histamine, PGE, nitric oxide (NO), and leukotrienes, which promote the cascade of inflammation [
64], resulting in the development of debilitating diseases such as RA and OA [
65]. Much research has been conducted to reveal the molecular mechanism of esculetin as a potent inhibitor of inflammation.
The transcription factor NF-κB is involved in the pathogenesis through the regulation of inflammatory genes [
66]. Briefly, the cytosol is a primary site of occurrence of NF-κB and under inflammatory stimuli; IκB is phosphorylated as well as degraded by the 26S proteasome resulting in the release of NF-κB. Consequently, the released NF-κB translocates to the nucleus, resulting in its binding to the promoter region and upregulation of inflammatory genes. A study by Yao-Jun et al. reported that esculetin at 20, 40 and 60 mg/kg downregulates the expressions of inflammatory cytokines and chemokines such as TNF-α, IL-1β, IL-6, CCL2, and inducible nitric oxide synthetase (iNOS). They also highlighted that the same dose of esculetin inhibits the NF-κB, STAT1, STAT3 and p65 expressions in LPS-induced macrophages and septic mice. In addition, the translocation of p65 from the cytoplasm to the nucleus was also inhibited upon esculetin treatment in macrophages. The phosphorylation of IKKα/β, IKBα, ERK1/2, JNK, and JMJD3 was also observed to downregulate in esculetin-treated LPS-induced macrophage cells [
67]. Esculetin with a 12 µg/mL dose potentially inhibits the TNF-α, IL-1β, ROS, and LPS-mediated nuclear translocation of NF-κB p65 by suppressing the IKβ-degradation in RAW 264.7 cells [
68]. A study by Jayakumar et al. found that esculetin reverse LTA-induced IkB degradation and NF-κB p65 phosphorylation. By increasing Nrf2 and scavenging DPPH radicals in RAW 264.7 cells at a concentration of 20 µM, esculetin inhibited the translocation of NF-κB p65 to the nucleus [
69].
NO is one of the key mediators of inflammation [
70]. Nitric oxide synthase (NOS) is an enzyme that exists in three different forms: neuronal NOS (nNOS), endothelial NOS (eNOS), and iNOS. This class of enzyme is responsible for the conversion of L-arginine into NO [
71]. But when there is an imbalance in this reaction, NO levels rise, resulting in exacerbated inflammation. A study by Zhu et al. reported that 20 and 40 mg/kg doses of esculetin result in the inhibition of TNF-α, IL-1β, and IL-6 expressions in serum and hippocampus. Further, the downregulation of cyclooxygenase (COX)-2 and iNOS was also observed when treating the mice with esculetin. In addition, this coumarin compound with the same dose was also found to inhibit the LPS-induced pIKK-α, pIKK-β, p-IKB-α, and p-NF-κB65 activation. Esculetin also results in the activation of brain-derived neurotrophic factor/tropomyosin receptor kinase B (BDNF/TrKB) signaling pathway, by which the levels of p-TrKB protein were upregulated in serum and hippocampus, and thus, esculetin exhibits the neuroprotective activity [
72]. Similarly, the treatment of esculetin at 80 and 120 µM to RAW 264.7 cells and BALB/c mice results in an increase in endocytic activity and augmented NO and iNOS levels in LPS-treated macrophages [
73]. At 100 mg/kg and 200 mg/kg, esculetin inhibits the inflammatory cascade by attenuating the NO and PGE2 levels in synovial fluid and MMP-1 expression in the cartilage of osteoarthritic rabbits [
74].
In the adipose tissues of obese mice, macrophages are known to release potent inflammatory agents, especially NO, PGE, and TNF-α causing systemic inflammation. Administration of a 100 µM dose of esculetin upregulates the level of heme oxygenase 1 in cocultured macrophages and adipocytes by restricting the liberation of TNF-α, NO and MCP-1. Moreover, the inhibition in peroxisome proliferator-activated receptor (PPAR)-ϒ and CCAAT/enhancer binding protein α was also reported to inhibit with esculetin treatment, suggesting its protective role against the obesity-induced inflammation [
75]. Proinflammatory cytokines such as TNF-α, IL-1β, IL-2, interferon (IFN)-γ, IL-8, and IL-6 are produced during macrophage activation and leads to the inflammatory diseases [
76]. Both doses of esculetin (100 µM and 5 mg/kg) were found to inhibit the production of TNF-α, IL-1β, IL-2, INF-ϒ in rats and RAW 264.7 cells. Esculetin was also reported to prevent the generation of ROS and reduced glutathione (GSH) depletion. Similarly, the activities of myeloperoxidase (MPO) and ALP were inhibited with esculetin treatment [
77].
Esculetin at 5 µM concentration effectively reduces the LPS-induced phosphorylation of ERK1/2 and NF-κB expressions and protects the cells from apoptosis and necrosis as well. Moreover, reduction in levels of LPS-induced TRAIL, TNFR, IL-1β, IL-6, IL-12, vascular epidermal growth factor (VEGF), Manganese superoxide dismutase (MnSOD), and glutathione peroxidase (GPx) was observed with esculetin treatment in LPS-induced inflammation in human retinal pigment epithelial cells (ARPE-19 cells) [
81]. Both doses of esculetin (20 and 40 mg/kg, 0.1, 1, and 10 µM) were reported to inhibit the MPO activity, neutrophil infiltration, NF-κB pathway, RhoA/Rho kinase pathway, followed by the downregulation of TNF-α, IL-1β, and IL-6 in A549 cells and BALB/c mice [
82]. The TNBS-induced colitis in HCT116 cells and Sprague Dawley colitic rats was found to ameliorate with esculetin treatment at 100 and 200 µM doses. It inhibits MPO, COX-2, and iNOS expression and activates the HIF-1 in HCT116 cells, which results in the activation of hypoxia-inducible factor (HIF)-1α and VEGF. Furthermore, esculetin was also reported to prevent the activation of HIF-prolyl hydroxylase-2enzyme and thus exhibits anti-colitic effects [
83].
A study by Chen et al. highlighted the anti-psoriatic effects of esculetin in imiquimod-induced psoriasis in BALB/c mice. They showed that esculetin at 50 and 100 mg/kg doses significantly ameliorate the skin lesions by decreasing the expressions of TNF-α, IL-6, IL-22, IL-23, IL-17A, and IFN-γ cytokines. Moreover, repression of phospho-IKKα and phospho-p65 was also observed with the inhibition of the NF-κB signaling pathway. The reduction in effector CD8+ T cells and upregulation in CD4+ FOXp3+ Treg frequency in spleen and lymph nodes results in amelioration of skin disease with esculetin therapy. This coumarin compound decreased the Ki67 and K10 mRNA expressions and the CD3+ and CD8+ T-cell infiltration in psoriatic mice [
84] (
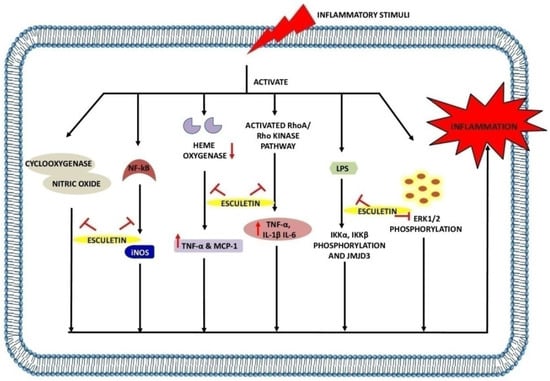
Table 3,
Figure 4).
Figure 4. Anti-inflammatory effects of esculetin: Esculetin inhibits the action of cyclooxygenase, NF-κB, NO, LPS, heme oxygenase, RhoA/Rho kinase pathway, and ERK1/2 phosphorylation, inhibiting the inflammatory reactions. ↑: Overexpression; ↓: low expression; ⊣: inhibition.
Table 3. Mechanism and function of esculetin in treatment of inflammation.
This entry is adapted from the peer-reviewed paper 10.3390/ijms232012643