Climate change and the associated disturbances have disrupted the relative stability of tree species composition in hemiboreal forests. The natural ecology of forest communities, including species occurrence and composition, forest structure, and food webs, have been affected. Yet, the hemiboreal forest zone of Lithuania is the least studied in the country for climate change risks and possible management adaption techniques. Moreover, the adverse effects of the phenomenon of global climate change can undermine the resilience of forest ecology in terms of its capacity for natural regeneration to occur successfully on the scale of the expected time. Especially since forests are multi-scale, multi-species networks that constantly evolve toward the successional processes and patterns of natural regeneration which cannot be reached at an individual tree species level. Therefore, in the context of hemiboreal forests, assisted natural regeneration lays the groundwork necessary to consider the life-cycle features of trees that affect the organic relationships between individual species and ecological communities indirectly via their effects on growth, reproduction, and survival, such as tree regeneration strategies that correspond to the various trade-offs in the adaptations to competition, stress, and forest disturbances. With the concept of assisted natural regeneration, embedded in the principle of deep ecology, the natural powers of forest ecology can be revitalized and fast-tracked to keep up with the pace of global climate change or even overtake it ahead its disruptions.
1. Introduction
Natural recurring forest processes are often self-organised and implicate sustainability processes in environmental changes. Niche construction, ecological engineering, and biosemiotics processes are different forms of indirect, background interaction and communication of organisms in the environment [
1]. Self-organisation of an ecosystem includes all the diversity that cannot be reduced to the properties of an individual system’s components, such as molecules, genes, populations, and species in both time and space [
2]. Meaning is generated across all the organisational levels [
3]. The strategy of life expansion is realized through the spread of life in space—the proliferation and collaborative construction of ecosystems and the biosphere by organisms. Forests are multi-scale, multi-species networks that constantly evolve toward the successional processes and patterns of natural regeneration which cannot be reached at an individual tree species level. In this direction, our work focuses on the super-organism approach of forest communities that considers succession as a comprehensive ecological process of multiple events where the forest vegetation communities are directly related to environmental condition with regard to climate change [
4].
The life history traits of species are controlled by natural patterns and processes recurring over time and space at multiple scales [
5,
6]. Natural selection has matched trees to site and environmental conditions for millennia [
7] and is considered a key evolutionary process that can increase the adaptation rate of species to environmental change [
8]. Natural selection can be confirmed through field observations of ecological communities and their development towards self-organisation. Tree species’ life histories, reproductive character, regeneration times, mode of dispersion, and other evolutionary phenomena are interconnected in the immense and complex system of self-sustaining interactions of forest communities [
9]. The ecology of a forest never ceases to evolve. The probability of seed germination, tree growth, development and recruitment is dependent on a species’ genetic profiles and life history traits to cope with the changes in environmental conditions [
10,
11,
12]. Dynamics in forest communities are driven by a wide range of factors, including species’ invariable life history strategies [
4].
However, traditional forest management, climate change and increased disturbances have disrupted the relative stability of tree species composition in combination with the edaphic site conditions in European hemiboreal forests [
13,
14]. This is a key problem. The natural ecology of forest communities, including species occurrence and composition, forest structure, and food webs have been affected [
15]. Developing knowledge about natural forest disturbance dynamics and their relationship to anthropogenic impacts and management practices is essential towards the mitigation of impacts on forest ecosystems in the light of climate change [
16]. This warrants the basis for proposing the assisted natural regeneration (ANR) strategy as an alternative adaptive model for forest management in Europe.
2. European Hemiboreal Tree Species: The Case of Lithuania
We focused on the European hemiboreal forests of Lithuania because it is one of only two countries (Latvia and Lithuania) that falls completely within the hemiboreal forest zone in Europe [
17]. The hemiboreal forest zone is the flux zone between the temperate forest zone to the south and boreal zone to the north. Unfortunately, the forests of the hemiboreal zone are often overlooked in climate impact and adaption studies, while attention is focused on the other two zones [
14].
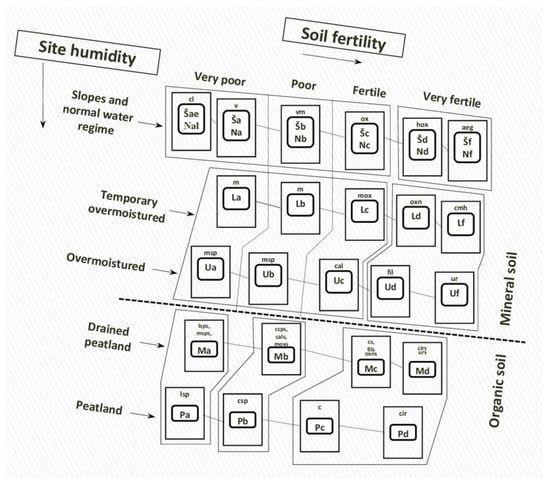
Lithuania’s hemiboreal forest site types (as well as the 13 Natura 2000 forest habitat types of European Community) can be classified into three main forest habitat types based on the concept of potential vegetation and soils [
14,
18,
19]: (1) mixed broadleaved forests on rich sites; (2) mixed species forests on mesic sites dominated by Norway spruce; and (3) Scots pine (
Pinus sylvestris) forests on poor sites. Soil moisture and fertility of Lithuania’s forests are considered the main drivers of forest disturbances and succession [
17]. As such, Lithuania’s forests have been classified by the Food and Agriculture Organisation (FAO, Rome, Italy) soil classification system [
20,
21] based on soil typological groups (
Figure 1).
Figure 1. Lithuania’s forest site type scheme. The bold codes refer to Lithuanian’s forest site types based on soil fertility and moisture, and the small non-bold letters refers to the forest vegetation type series [
18,
20,
23]: N—normally moist, L—temporarily over moist, U—over moist, P—peatland, and f—very eutrophic soils, d—eutrophic soils, c—mesotrophic soils, b—oligotrophic soils, a—very oligotrophic soils; aeg—
Aegopodiosa, c—
Caricosa, cal—
Calamagrostidosa, cir—
Carico-iridosa, cl—
Cladoniosa, cmh—
Carico-mixtoherbosa, csp—
Carico-sphagnosa, fil—
Filipendulo-mixtoherbosa, hox—
Hepatico-oxalidosa, lsp—
Ledo-sphagnosa, m—
Myrtillosa, mox—
Myrtillo-oxalidosa, msp—
Myrtillo-sphagnosa, ox—
Oxalidosa, oxn—
Oxalido-nemorosa, ur—
Urticosa, v—
Vacciniosa, vm—
Vaccinio-myrtillosa.
3. Hemiboreal Tree Dynamics
3.1. Tree Regeneration Strategy
Forests are characterized by the development of contiguous communities of trees that are relatively uniform in composition, structure, age, size, class, distribution, spatial arrangement, site quality, condition, and location to distinguish them from adjacent communities created by human intervention [
24,
25,
26]. The absence of structural legacies at multiple scales is one of the most distinguishing features of modified forests subjected to intense and frequent anthropogenic disturbances [
27,
28]. Species’ life history traits are interrelated with natural disturbances and associated site conditions, and these account for the interactions (patterns and processes) in species distribution [
14,
29]. There is also increasing evidence that the intrinsic influences of disturbance susceptibility are phylogenetically inherited, implying that species-level traits are constrained by developmental, genetic, or other correlated limitations [
30]. Being the primary species of forest ecosystems, long-lived trees are pivotal in providing associated organisms with a combination of resources and habitats that range from beneficial to detrimental [
31]. Therefore, the forest development and growth dynamics of tree species follow relatively fixed patterns and can be difficult to modify in the light of the interactions of both biological and physical processes. This is also the case with hemiboreal trees’ natural regeneration.
There are four tree natural regeneration strategies, i.e., the establishment and growth of trees in forest gaps [
32,
33,
34,
35,
36]: (i) colonization; (ii) occupation; (iii) invasion; and (iv) expansion (
Table 1). These are inter-intuitive with Clark and Clark’s [
37] tree species regeneration groups (A–D), Whitmore’s [
38] tree species groups, having an increasing “pioneer index” (1–4), and Grime’s [
39] four types of secondary ecological strategies in trees that are derived from the theoretical triangular scheme of competitor (C), stress-tolerant (S) and ruderal (R) primary plant ecological strategies—stress-tolerant ruderals (S-R), competitive stress-tolerant ruderals (C-S-R), competitive ruderals (C-R), and competitive stress-tolerators (C-S). Colonization (D, 4, S-R) implies that even-aged seedlings are being established after gap formation and grow only in gaps. This relates to stress-tolerant species that possess a ruderal strategy without advanced regeneration. Juveniles have the highest growth potential. A ruderal strategy is a characteristic of many species that never become established in ruderal habitats. Ruderal species are plants that grow only in habitats that have been completely disturbed and damaged by human activity [
40]. Occupation (C, 3, C-S-R) relates to the competitive stress-tolerant ruderal strategy species occurring as gap makers. Their seeds germinate better in gaps with intermediate canopy openness than in the understorey or large gaps, saplings can survive in closed forests. Invasion (B, 2, C-R) implies that trees regenerate from saplings recruited before gap or stand formation. This type involves competitive species with a ruderal strategy of advance regeneration, allowing already established juveniles to survive in newly created gaps. Expansion (A, 1, C-S) implies that trees in the forest regenerate as advanced regeneration under shade. This usually involve competitive stress-tolerant species. Juveniles have average growth rates.
Table 1. Conceptual framework of hemiboreal tree dynamics in Lithuania [
32,
48,
50]. Capital letters indicate the main tree species that form forest stands in gap dynamics (G), successional development (S), or multi-cohort succession (M), whereas small letters (g, s, and m) indicate secondary ones which are a valuable admixture in these stands.
3.2. Natural Regeneration of European Hemiboreal Tree Species
Multiscale recovery dynamics analysis of community typology is measured to determine the impact of change in forest ecology. Usually, it ranges from tree genetic variation characteristics (in terms of regeneration vs. canopy compositions) to multi-population structures reflected in disturbance and management regimes. To enhance the adaptive potential and associated ecosystem services of forests, we proposed a conceptual framework for hemiboreal tree dynamics based on a dynamic typology of forest communities [
13,
14,
18,
19,
41,
42,
43,
44,
45,
46] and forest sites defined by field layer-canopy dominants, on-site soil fertility and moisture [
18,
47], and four types of tree regeneration strategies [
33,
34,
35,
37,
38,
39,
48] (
Table 1). It follows the Lithuanian classification of forest types and the layer dominants: forest site type, forest type series (field flora), and dominant and secondary tree species [
18]. The three dynamic forest habitat types in our conceptual framework represent general descriptions of plant community types that reflect the dynamics of vegetation cover that occur in the course of natural disturbances [
13]. In hemiboreal forests, there are three main types of natural disturbance regimes that determine the success of natural selection: (1) gap dynamics caused by the death of individual trees or small groups of trees in the absence of fire; (2) successional development after severe stand-replacing disturbances, such as crown fires, large windthrows, pest outbreaks, etc.; and (3) multi-cohort dynamics related to partial disturbances, such as low-intensity surface fires [
41,
42,
43,
44,
45,
46].
Hemiboreal forests may be legacies of biological and physical disturbances [
6,
24]. Disturbance regimes are classified by the type, magnitude and duration of environmental variation as well as community (ecosystem) and individual species resilience [
42,
43,
44,
49]. Tree species regeneration in hemiboreal zone is generally rapid after large-scale short-term disturbances (e.g., forest fire) but slower after longer term disturbances such as repeated logging or forest conversion to monoculture plantations [
48]. Restoration of the original forest ecosystem via natural regeneration can take several centuries as succession begins with early-successional herb, shrub, and tree species, and culminates with late-successional species. In order to understand forest regeneration processes following a disturbance, one needs to be knowledgeable in forest dynamic typology, which can provide a first insight into the status of vegetation cover (i.e., basal, canopy, foliar, or ground cover) and warn us if it is facing decline or an unwanted trajectory. As such, we have allocated each hemiboreal tree species to a dominant regeneration strategy (
Table 1).
However, a forest stand that is subject to a larger-scale disturbance can also be subject to smaller-scale disturbance thus the scale of disturbance is also a factor that needs be considered and discussed. For instance, in mixed Norway spruce forest, disturbance can range from a single tree (gap/small patch (G)) to a stand or forest (large patch (S)) sized disturbance. Thus, there is no certain rule on regeneration. Another factor to consider is the type of disturbance. For instance, Scots pine is fire tolerant, and fire stimulates regeneration. Conversely, Norway spruce is fire intolerant and thus is often eliminated together with its seed bank. So, fire creates multi-cohort pine stands and eliminates spruce. Also, the different disturbance regimes of forests undergo generate different age profiles [
27].
This entry is adapted from the peer-reviewed paper 10.3390/d14100892