Stem cells are a well-known autologous pluripotent cell source, having excellent potential to develop into specialized cells, such as brain, skin, and bone marrow cells. The oral cavity is reported to be a rich source of multiple types of oral stem cells, including the dental pulp, mucosal soft tissues, periodontal ligament, and apical papilla. Oral stem cells were useful for both the regeneration of soft tissue components in the dental pulp and mineralized structure regeneration, such as bone or dentin, and can be a viable substitute for traditionally used bone marrow stem cells. The role of bioactive compounds in oral stem cell-meditated regeneration is discussed.
- oral stem cells
- ginsenoside Rg1
- apigenin
1. Oral Stem Cell-Mediated Dental Regeneration
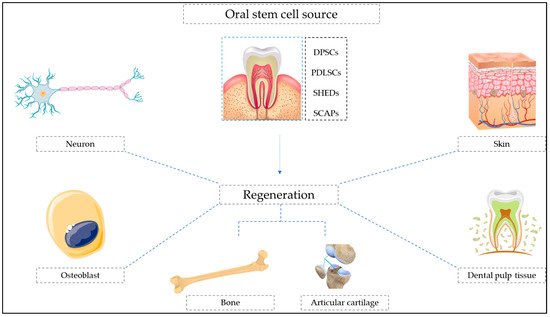
2. Botanicals in Oral Stem Cell-Mediated Regeneration
| Source | Bioactive Compounds | Type of Study | Major Findings and Mechanism of Action | References |
|---|---|---|---|---|
| Artemisia annua | Artemisinin (Sigma Aldrich, St. Louis, MO, USA) |
In vitro study investigated the effect of Artemisinin on hypoxia and TNF-α mediated osteogenesis impairment in DPSCs | Artemisinin reversed the suppression of cell survival caused by hypoxia or inflammation in DPSCs, along with restoring the osteogenic differentiation potential of DPSCs | [91] |
| Sapindus mukorossi | Seed oil (He He Co., Ltd., Taipei, Taiwan) |
In vitro study to examine the effects of S. mukorossi (seed oil) on the differentiation and proliferation of DPSCs | Enhanced the odontogenic/osteogenic differentiation potential of DPSCs by upregulation of ALP gene expression and mineralization-related extracellular vesicle secretion | [175] |
| Curcuma longa | Curcumin (Sigma Aldrich, St. Louis, MO, USA) |
In vivo study on effect of curcumin on hPDLSCs osteogenic differentiation | Curcumin increased protein and mRNA levels of COL1, ALP, RUNX2, and activated PI3K/AKT/Nrf2 signaling pathway | [185] |
| Curcuma longa | Curcumin (Solarbio Life Sciences, China) |
In vivo study Curcumin displays promoting osteogenic differentiation and its mechanism |
Curcumin 10 µmol/L treatment maximal promoting the cells viability, ALP activities, mineralization, and levels of Runx2, OC, OPN, Collagen I, and EGR-1 in hPDLSCs |
[186] |
| Berberis vulgaris | Berberine hydrochloride (Wako Pure Chemical Industries, Ltd., USA) |
In vitro study to examine effects of AGE and berberine hydrochloride on the hPDLSCs’ osteogenic differentiation ability | Berberine hydrochloride was able to reverse the inhibition of the PDLSCs’ osteogenic potential in an AGEs enriched microenvironment, partly by inhibition of the β-catenin and canonical Wnt pathway | [187] |
| Berberis vulgaris | Berberine (Sigma Aldrich, St. Louis, MO, USA) |
In vivo study to examine the effect of berberine on rat root canals of immature teeth with apical periodontitis | Berberine induced β-catenin expression and activated the β-catenin and canonical Wnt pathway in SCAPs which improved root repair in immature teeth with apical periodontitis. |
[188] |
| Berberis vulgaris | Berberine (Sigma Aldrich, St. Louis, MO, USA) |
In vitro study to examine effects of berberine on the osteogenesis and cell proliferation of DPSCs | Berberine enhanced hDPSC cell proliferation in a dose-dependent pattern and activated MAPK–EGFR–Runx2 signaling pathways. | [189] |
| Reynoutria multiflora (Thunb.) Moldenke | 2,3,5,4′-Tetrahydroxystilbene-2-O-β-D–glucoside (THSG) (Taipei Medical University, Taipei, Taiwan) |
In vitro study investigated the effect of THSG on cell proliferation in hDPSCs. | THSG treatment enhance d the renewal ability and proliferative potential of hDPSCs via the AMPK/ERK/SIRT1 axis |
[191] |
| Dictamnus dasycarpus | Fraxinellone (Chengdu Herbpurify Co., Ltd., Chengdu, China) |
In vitro and in vivo study to examine antitumor effects of fraxinellone on lung cancer cells | Fraxinellone treatment inhibits expression of PD-L1 by HIF-1α and STAT3 signaling pathway downregulation, further inhibiting angiogenesis and proliferation in cancer cells | [194] |
| Fagopyrum esculentum | Rutin (Solarbio Science & Technology Co., Ltd., Beijing, China) |
In vitro study to examine the effects of rutin on the PDLSCs’ osteogenic proliferation and differentiation | Rutin increased osteogenic differentiation and proliferation of PDLSCs by GPR30-mediated PI3K/AKT/mTOR signal transduction | [195] |
| Cnidium monnieri | Osthole (National Institutes for Food and Drug Control, Beijing, China) |
In vitro study to determine osthole efficiency against defective osteogenic differentiation of P-PDLSCs via epigenetic modification |
Osthole (10−7 Mol/L) upregulated MORF, MOZ, and histone acetylases that catalyze acetylation of Histone 3 lisine14 (H3K14) and Histone 3 lisine9 (H3K9) | [199] |
| Osthole treatment enhanced bone formation and cell sheet formation of PDLSC sheets in periodontitis (animal models) | ||||
| Drynaria roosii Nakaike | Naringin (National Institute for the Control of Pharmaceutical and Biological Products, China) |
In vitro and in vivo study to examine the effect of naringin on proliferation and osteogenic differentiation of hPDLSCs | Naringin (1 µM) efficiently promoted hPDLSC differentiation and proliferation and increased expression levels was observed in related genes (COL1A2, OPN, RUNX2, and OCN) as compared to the control group | [201] |
| Macaranga tanarius | Isonymphaeol B (Okinawa, Japan) |
In vitro study to identify odontogenesis-promoting compounds and examine the molecular mechanism underlying enhanced tooth formation and odontoblast differentiation | Isonymphaeol B shows stimulatory effects on tooth root, dentine formation and odontoblast differentiation via AKT and MAP kinase signaling pathways | [203] |
| Canavalia ensiformis | Concanavalin A (Sigma Aldrich, USA) |
In vitro study to determine the effect of concanavalin A on osteogenic and proliferation differentiation of DPSCs | Concanavalin A at concentration of 5 and 10 µg/mL to DPSCs significantly increased the osteogenic and proliferation differentiation of DPSCs (p ≤ 0.05) | [205] |
| Panax ginseng | Ginsenoside Rg1 (Bio-function, Beijing, China) |
In vitro study to examine the effects of ginsenoside Rg-1 on osteogenic differentiation and proliferation of hPDLSCs | Ginsenoside Rg-1 enhanced osteogenic differentiation and proliferation of hPDLSCs at an optimum concentration of 10 μmol/L | [209] |
| Eclipta prostrata | Wedelolactone (Dalian, China) |
In vitro study investigated effect of wedelolactone on odontoblast differentiation of DPSCs | Wedelolactone induced odontoblast differentiation through NRP1, Sema3A, and NF-κB pathway inhibition and activation of b-catenin pathway | [210] |
| Aloe barbadensis Mill. | Acemannan (Chulalongkorn University, Bangkok, Thailand) |
In vitro study to examine effects of acemannan on human deciduous pulp cells and the response after vital pulp therapy in dog deciduous teeth | DPSC proliferation was significantly enhanced by acemannan along with an increase in expression levels of type I collagen, BMP-2, vascular endothelial growth factor, BMP-4, alkaline phosphatase, dentin sialoprotein, and mineralization | [211] |
| Cirsium setidens (Dunn) Nakai | Methanolic extract (Kangwon National University, Republic of Korea) |
In vitro study to examine the effects of methanolic extracts of C. setidens on osteogenic differentiation of hPDLSCs | Methanolic extract treatment with concentration of 0.05% significantly increased the viability of PDLSCs and also increased the expression levels of alkaline phosphatase, collagen 1, runt related transcription factor 2, and bone sialoprotein | [218] |
| Rhodiola rosea | Salidroside (Chengdu Must Bio-Technology Co., Shanghai, China) | In vitro study to investigate the effect of salidroside on the odontogenic differentiation and proliferation of hDPSCs | Treatment with salidroside promoted DPSCs cell viability, along with promoting their differentiation into odontogenic and osteogenic linage via activation of the BMP signaling pathway | [219] |
| Moringa oleifera | Moringin (Indena India Pvt. Ltd.; Bangalore, India) |
In vitro study to examine efficacy of moringin to induce PDLSCs toward neural progenitor differentiation | Treatment of PDLSCs with moringin resulted in the induction of PDLSC differentiation to neural progenitor cells via increased gene expression levels of genes that were involved in neuron cortical development | [224] |
| Acacia nilotica | Aqueous leaves extract (Hormavu, Bangalore) |
In vitro study to investigate the efficacy of Acacia nilotica leaves extract in chondrogenesis induction from mesenchymal stem cells | Treatment of aqueous leaves extract promoted chondrogenesis induction in DPSCs by upregulating the expression of various proteins in the cellular matrix, such as aggrecan, sox9, glycosaminoglycan (GAG), and collagen 2α1 (Col2α1) | [231] |
| Acalypha indica | Methanolic extract (Porur, Chennai, India) | In vitro study on GMSCs highlighted the potential of A. indica (methanolic extract) in increasing the wound healing ability of GMSCs | Treatment of A. indica extract (25 µg/mL) wound closure activity of GMSCs was increased up to 56.91 ± 1.21% in 24 h, while the percentage of wound closure was further enhanced to 89.23 ± 1.09% post 48 h of treatment | [232] |
| Scutellaria baicalensis | Baicalein (Sigma Aldrich, USA) |
In vitro study investigated the effect of Baicalein on Angiogenesis and Odontoblastic Differentiation | Baicalein promoted odontoblastic differentiation and angiogenesis of HDPCs by activating the BMP and Wnt/β-catenin signal pathways | [233] |
| Salvia miltiorrhiza | Tanshinone IIA (Sigma Aldrich, USA) |
TSA affects the osteogenic differentiation of hPDLSCs. | Tanshinone IIA can induce hPDLSC osteogenesis through the ERK1/2-Runx2 axis | [236] |
| Cullen corylifolium L. | Bakuchiol and C. corylifolium extract (KMD medicinal herbs Co., Yunnan, China) |
In vitro study to examine the efficacy of Bakuchiol and C. corylifolium extract as differentiation-inducing substances for tooth regeneration, was determined by monitoring odontogenic differentiation in hDPSCs | Bakuchiol and C. corylifolium extract significantly enhance the odontogenic differentiation potential of hDPSCs via upregulation of odontogenic differentiation marker genes, such as dentin matrix acidic phospho-protein-1, alkaline phosphatase, osteocalcin, and Runt-related transcription factor 2 | [237] |
This entry is adapted from the peer-reviewed paper 10.3390/cells11182792