Taro [Colocasia esculenta (L.) Schott] is the most commonly cultivated species in genus Colocasia and is the fourth most consumed tuber crop globally. It is a member of family Araceae, sub-family Aroideae, and is a tropical monocotyledonous, vegetatively propagated, perennial crop grown primarily for its starchy corm or underground stem. Taro is one of the world’s oldest food crops, with its domestication dating back over 9000 years. It was probably first domesticated in Southeast Asia and thereafter spread across the world, to become one of the most important staple food crops in the Pacific Islands. It is widely distributed across Africa, Oceania, Asia, and the Americas [4,5]. The crop has been largely maintained by smallholder farmers, and the species’ genetic resources have remained largely within local communities. In many societies, taro is considered a sacred plant of strong cultural importance and is used in religious festivals, domestic and agricultural rituals, and as bride price.
1. Taro Production in Africa
The current distribution of taro as a cultivated food crop extends from southern to northern Africa [
6,
33], western Asia to eastern Asia, across Southeast Asia and the Pacific Islands, and through the Americas, from the USA to Brazil [
34]. The root crops that have an important role in many African countries are potato, cassava, sweet potato, yam, taro, and the new cocoyam [
35]. Within Africa, four countries, Nigeria, Ethiopia, Ghana, and Cameroon, accounted for about 67% of total production in 2020 [
16]. Numerous taro cultivars are found across WA and occupy an important role in local agriculture and traditions, indicating a long history of the crop in the region. Grimaldi [
36] produced a taro distribution map for the period between 1849 and 2012 based on several reports and academic articles on taro cultivation at specific locations and region in African countries.
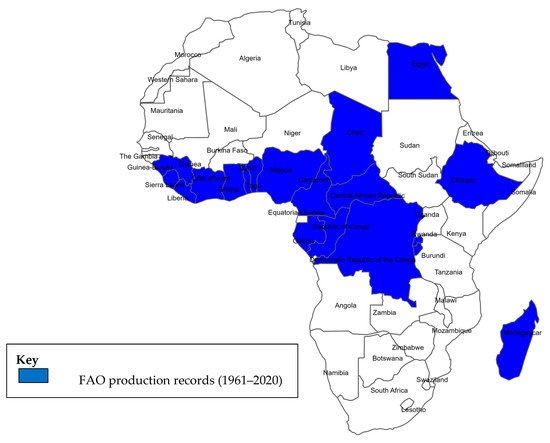
Figure 1 shows the distribution of African countries where taro was cultivated between 1961 and 2020, indicating that it is grown in hot, humid areas with high rainfall, such as the tropical sub-Saharan region, but that it can also be grown in drier regions along streams, such as in Egypt, Algeria, and Libya. Taro can thrive under diverse agro-ecological and soil conditions and is referred to as an ecologically friendly crop [
37]. The crop optimally grows at altitudes extending from 60 to 1850 m above sea level in the tropics and temperate zones [
7,
38,
39]. However, despite its wide geographical presence, there are limited research effort and funding for the large-scale assessment of the production, trade, and usage of this crop. Changes in the environment and agricultural systems in Africa have also led to a decline in taro production.
Figure 1. Map of taro production in Africa based on FAOSTAT data from 1961 to 2020.
Traditionally, taro is propagated using corms, cormels, suckers, and tops (huli, a Hawaiian vernacular term used to describe a plant part). The use of tops and suckers is preferred because the growing season from planting to harvest is shorter than that of cormels, corms, or corm pieces (setts), although cultivars with many suckers produce smaller corms [
37]. The yield of taro is optimized when the soil is appropriately managed and agronomic practices are carried out well [
3]. On average, taro requires a well-drained sandy loam soil with good water retention capacity, and benefits from the application of NPK fertilizers [
18,
40]. Quality organic materials and bio-fertilizers (Mycorrhizae, Azotobacter, and Phosphorine) can also be used via furrow placement and split application, respectively [
3].
2. Taro Production Constraints in Africa
The major biotic stress for taro is TLB, a disease caused by
P. colocasiae; it is an oomycete disease with highly devastating effects [
44].
Phytophthora colocasiae was first reported in Java by Raciborski [
45] and has now spread all over the tropics [
46,
47]. The disease thrives where day and night temperatures range between 25–28 °C and 20–22 °C, respectively, and can assume epidemic proportions all year round under favorable conditions [
48]. TLB was not known in WA before 2009, when there were simultaneous outbreaks in Nigeria, Cameroon, Ghana, and other neighboring countries [
20,
21]. It is estimated that TLB in WA accounts for an economic loss of about USD 1.4 billion annually [
19].
Phytophthora colocasiae reproduces asexually during rainy seasons, with the production of sporangia from sporangiophores at the extremity of lesions in infected leaves. Sporangia leave the pedicel during rain fall and germinate to produce motile zoospores that can swim for short distances in water and encyst to form germ tubes that can penetrate the host. This can happen within two hours at a favorable temperature of 20 °C and a minimum humidity of 90%. At an ideal temperature of 24–27 °C, symptoms present 2–4 days after penetration of the germ tube [
46,
47].
Phytophthora colocasiae is heterothallic with two mating types, A1 and A2, and can produce oospores via sexual reproduction [
49].
In the field,
P. colocasiae is spread mainly by zoospores and sporangia. The propagules are short-lived in the infected leaves and tissues and are carried by water to a host through rain splashes [
47,
50]. Taro corms are, however, rarely harvested from the field and can sustain the pathogen. Usually, planting is carried out within a short time frame after harvesting; any infected tissue debris left in the field is, therefore, a source of inoculum for subsequent infection of new plants [
49,
50]. An entire field can become symptomatic of the disease within seven days when conditions are favorable [
19,
48]. A symptomatic plant initially displays water-soaked lesions with a diameter of 1.5 cm around the leaf edges; the fluid exudates from the lesions are of bright-yellow to dark-purple colors when dried. As the disease progresses, the lesions enlarge, developing a zonate appearance characterized by brownish to purplish-brown colors. White fuzz also appears on both sides of the leaves, indicating sporangia, which continues to develop until the leaves are completely covered [
46,
48]. The white fuzz of sporangia around lesions is a characteristic symptom of
P. colocasiae infection [
46]. The infected leaf tissues collapse after 20 days, unlike healthy leaves, which last 40 days before senescence [
50]. TLB also affects the corms, causing them to rot, and yield losses as a result of this disease can be as high as 70–100% [
46,
48,
51].
TLB is usually controlled with the use of copper fungicides at a rate of 38 Lha
−1, using 2.24 kg of copper oxychloride as the active ingredient. Fungicide applications start from four months after planting (MAPs) and continue until nine MAPs, weekly during rainfall periods and bi-weekly when conditions are dry. Dithane M-45 can also be used, at a rate of 1.68–2.25 kilograms in 189.3–378.5 liters of water per hectare. This application can be weekly or bi-weekly, depending on the severity of the disease, but should not exceed 25 applications and cannot be used once the crop is nine months old [
48]. Metalaxyl fungicides have also been effectively used to control TLB [
49]. In Hawaii, planting distance has been used as a control measure, with a decrease in disease incidence achieved by increasing the planting distance from 46 cm to 75 cm. In the Solomon Islands, improved sanitation, via pruning and removing infected leaves on a bi-weekly basis, has also reduced disease incidence.
The use of resistant and immune varieties is the most viable control measure for TLB in terms of environmental impact and sustainability [
43,
52,
53,
54,
55,
56,
57]. However, the use of resistant varieties has been limited by a lack of crop improvement programs and a lack of desirable economic market value traits in resistant genotypes. This is compounded by a lack of understanding of the genetic structure of pathogen populations [
56]. Compared with other species of
Phytophthora, very little attention has been paid to
P. colocasiae, either globally or at a regional level. Some research has been carried out on screening for disease-resistance genotypes and their adaptability in WA and beyond. For example, Ackah et al. [
58] evaluated taro genotypes from Ghana for resistance to TLB and found all the genotypes to be susceptible to varying degrees. Similarly, Amadi et al. [
58] characterized some local and exotic collections of taro for yield, local adaptation, and TLB resistance in Nigeria and found some promising genotypes, although no single genotype combined all the desired traits.
Additional major taro diseases are caused by viruses and other microorganisms that are specific to the Pacific [
41]. These reduce corm size and quality, with yield losses of up to 20%. For example, the co-infection of taro with taro bacilliform virus (TaBV) and Colocasia bobone disease virus (CBDV) is thought to be lethal to the crop. TaBV, along with taro bacilliform CH virus (TaBCHV) diseases [
59] and dasheen mosaic virus disease [
10], has been reported in Africa. Several other taro viruses have also been found in the Pacific, which currently restricts international movement of germplasm; thus, many countries do not have access to agronomically elite genotypes and selected traditional cultivars. Taro diseases reported in the Pacific include taro soft rot, caused by several species of
Pythium, sclerotium rot, caused by
Sclerotium rolfsii, and cladosporium leaf spot, caused by
Cladosporium colocasiae [
41]. Taro soft root rot and cladosporium leaf spot have been reported in Africa [
19,
60].
3. Opportunities for Improved Taro Production in WA
Among root crops, taro currently has the lowest average yield (5.4 tons/ha globally) [
92]. WA countries dominate as the major producers of taro globally, producing 4.9 million tons of the estimated 12.0 million tons of taro produced in the world [
79]. In the same year, Nigeria, the world’s largest producer of taro, harvested approximately 3.2 million tons from 0.8 million hectares, followed by Ghana with 1.68 million tons [
81]. Currently, there is no taro germplasm repository in WA responsible for preserving, characterizing, and distributing taro germplasm. The International Institute of Tropical Agriculture (IITA) in collaboration with the National Root Crop Research Institute of Nigeria (NRCRI) had collected and preserved taro landraces. However, this collection was lost during the outbreak of TLB in West and Central Africa, including Nigeria [
20], Ghana [
21], and Cameroon [
93]. Breeding resistant cultivars offers the best long-term control of TLB disease in most production systems; thus, urgent and collaborative efforts among research groups and donors are needed to combat the TLB epidemic and prevent taro from going extinct in the region.
In recent years, scientists in WA and beyond have been gathering information, developing strategies, and evaluating stress factors to help to improve the taro crop [
8,
18,
19,
58,
64,
94,
95,
96,
97,
98]. Two of the improvements achieved to date are outlined as follows:
- i.
-
Standardized collection protocols: Dansi [
95] has outlined a collection procedure for root and tuber crops, including taro. The methodology is based on synthesized information from the publications of international bodies such as Biodiversity International, World Conservation Union (IUCN), FAO, and United Nations Environment Programme (UNEP). This helps to ensure that collection programs are executed using international standard procedures;
- ii.
-
Characterization of local and exotic germplasm for agro-morphological traits, disease resistance, and nutritional qualities, to be used as the basis for taro improvement: Several authors have used morphological characterization to evaluate taro cultivars, including agro-morphological traits, diseases, and flowering ability [
58,
75,
90,
97]. This has helped the classification of taro into different morphotypes, which can then be used in taro improvement programs.
There is still more to be done with taro in WA, however. There is a scarcity of information on the use of molecular techniques in the characterization of taro, which would facilitate the understanding of the genetic phylogeny of local accessions and fast-track the improvement of the crop. In addition, there is a need to raise awareness among different stakeholders, including producers and consumers, about the crop’s potential contribution to food security, health, and economics, so that the improved production of the crop is prioritized at regional and country levels.
4. Breeding Efforts in Taro
Taro breeding efforts should focus on traits that are important for both producers and consumers, such as yield, pest and disease resistance, nutritional quality, shelf life, plant architecture, maturity, and culinary characteristics [
99]. Breeding programs involving different stakeholders can be used to gather more information and adopt new technologies. For example, there is a need to assess the acridity levels of different cultivars so that consumers can develop suitable cooking methods for increased edibility.
Taro breeding was initiated in the late 1970s, and varieties were released in Fiji (1978), Samoa (1982; 1996), Solomon Islands (1978; 1992), Papua New Guinea (1993), and India (1995) [
100,
101]. The first successful controlled hybridization of taro in Nigeria was reported in 2015 [
18]. Breeding schemes such as bi-parental crossing and recurrent selection were used at an early stage of plant development for traits such as taro corm flesh and corm fiber colors, which were correlated with the color of different petiole zones [
102]. There have been several efforts in Papua New Guinea towards resistance breeding, but one of the difficulties is getting rid of deleterious traits from wild types. Breeding programs have been initiated in Hawaii for agronomic traits, and pests and disease resistance, to develop improved varieties for the restaurant and landscape trades [
103].
The major objectives of taro breeding so far have been to improve plant architecture (such as optimal number of suckers, absence of stolons, optimal number of leaves, etc.), corm yield, resistance/tolerance to diseases such as TLB, and quality traits (such as dry-matter content, low levels of phenolic compounds causing irritation, etc.) [
104]. Breeding activities with parents from a diverse genetic base could result in improved targeted traits. However, taro is usually propagated through vegetative means, seldom through flowers; the flowers are protogynous, making conventional breeding methods difficult [
32]. Taro breeding has been initiated in many countries within the South Pacific under two major programs, TaroGen and TANSAO [
105]. These programs have focused on hybridization to develop new cultivars with higher yields, better taste, and improved resistance to TLB [
106,
107]. Hybridization in taro is promising, but it is labor intensive and lengthy in terms of the induction of flowering (although gibberellic acid has been used to induce flowering in taro [
18,
47,
75]), pollination, and seed harvesting. It takes 10 years or more from pollination for a new improved cultivar to be developed [
108]. There is a scarcity of reports on taro breeding in WA, and the majority of the work is limited to the agro-morphological characterization or evaluation of local landraces. Amadi et al. [
18] did achieve 109 crosses using 15 exotic taro cultivars and 4 local Nigerian cultivars, of which only 20 crosses (18.3%) were successful, with 9 crosses reaching maturity and producing seeds. The limited success with taro breeding can be linked to the weak institutional capacities of most national institutes engaged in breeding, coupled with a lack of genetic resources and funding for establishing a sustainable taro breeding program in WA. The work of existing national, regional, and international networks should also be consolidated to optimize breeding methodologies.
The application of biotechnological techniques for the disease-free clonal propagation of taro plants is another viable option. Tuia [
109] developed an efficient taro multiplication protocol and reported that it was possible to eliminate viruses using meristem cultures. There are also reports of somatic embryogenesis in taro [
110,
111,
112], but the regeneration rate is low. Similarly, Fukino et al. [
113] reported transformation in taro (
C. esculenta var.
antiquorum) calluses by particle bombardment, but only two putative transgenic plants were obtained. Transformation in
Colocasia esculenta var.
esculenta via microprojectile bombardment [
114] and
Agrobacterium tumefaciens [
115,
116] was also reported. These efforts show that using biotechnology to generate taro plantlets is a possibility, but there is a need to validate the vigor of plants in terms of growth rate, pest and disease resistance, and corm characteristics, in addition to extensive field trials to record the frequency of somaclonal variations [
112].
This entry is adapted from the peer-reviewed paper 10.3390/agronomy12092094