The ketogenic diet (KD), a low-carbohydrate and high-fat diet, has been widely prescribed in weight loss programs for metabolic abnormalities. Furthermore, research has investigated the effects of KD on the treatment of numerous diseases, including cardiovascular disease (CVD) and cancer, due to its role in promoting ketolysis, ketogenesis, and modifying many other metabolic pathways with potential favorable health effects.
- ketogenic diet
- Metabolism
- Physiological Effects
1. History
|
KD Type |
Macronutrient Proportion (% of Total Energy) |
General Characteristics |
||
|---|---|---|---|---|
|
Carbohydrate |
Fat |
Protein |
||
|
Classic ketogenic diet |
4 |
90 |
6 |
Developed for epilepsy treatment |
|
Medium-chain-triglyceride (MCT) ketogenic diet |
17 |
73 (30–60% MCT) |
10 |
MCT supplements should be incorporated into all meals and snacks |
|
The modified Atkins diet (MAD) |
5 (10–20 g/day) |
65 |
30 |
No restriction on energy content, fluid, or protein |
|
The modified ketogenic diet (MKD) |
5 (30 g/day) |
65–80 |
20–25 |
No restriction on energy |
|
Very low-calorie ketogenic diet (VLCKD) |
13 (usually <30 g/day) |
44 |
43 (1.2–1.5 g/kg of ideal body weight) |
Total energy intake of <800 kcal/day |
|
Ketogenic Mediterranean diet/modified Mediterranean ketogenic diet |
<30–50 g/day |
45–50 |
30–35 |
With an emphasis on lean meats, fish, olive oil, walnuts, and salad |
2. Classification
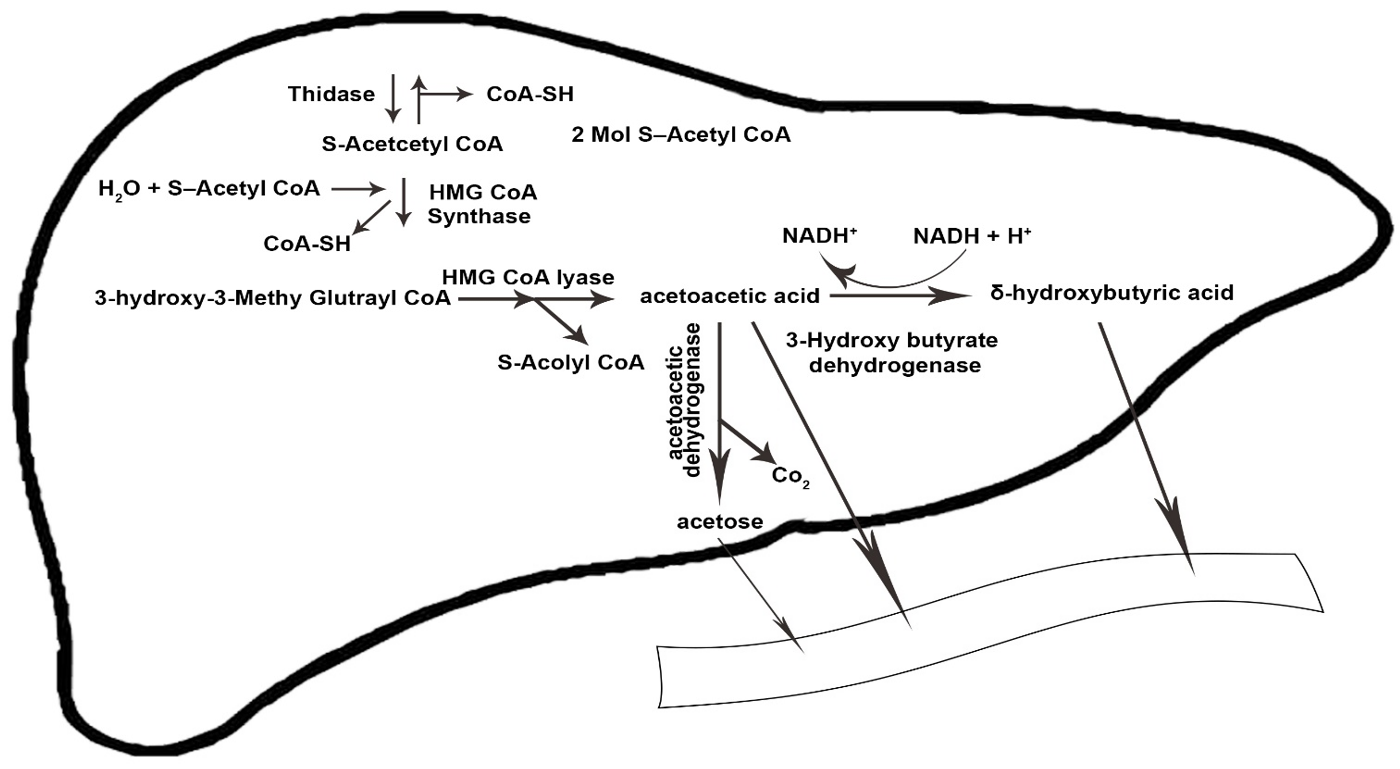
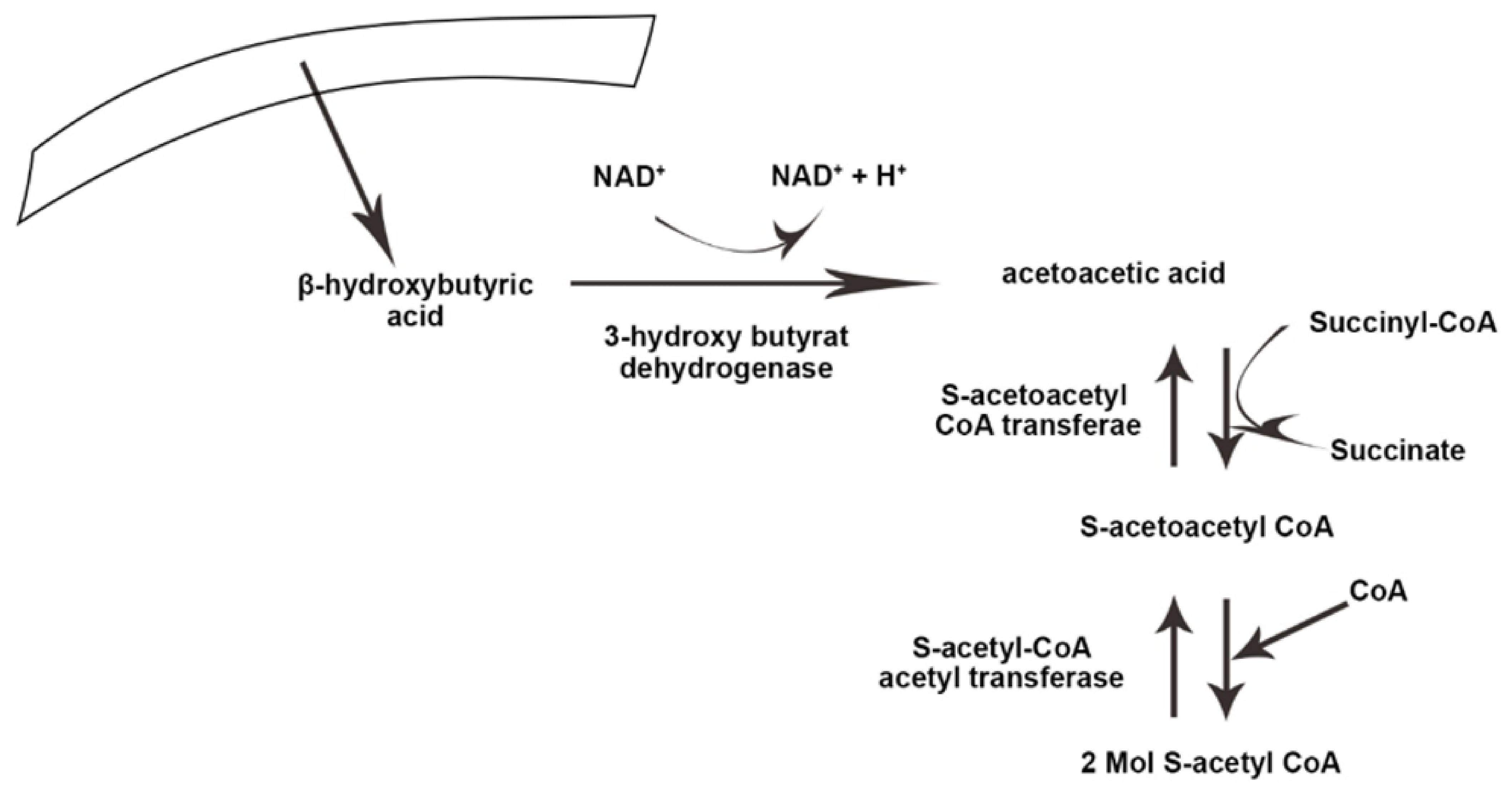
3. Physiology and Metabolism
This entry is adapted from the peer-reviewed paper 10.3390/nu14173499
References
- Williams, M.S.; Turos, E. The Chemistry of the Ketogenic Diet: Updates and Opportunities in Organic Synthesis. Int. J. Mol. Sci. 2021, 22, 5230.
- Guelpa, G. La lutte contre l’epiepsie par la desintoxication et par la reeducation alimentaire. Rev. Ther. Med. Chir. 1911, 78, 8.
- Wilder, R.M. The effects of ketonemia on the course of epilepsy. Mayo Clin. Proc. 1921, 2, 307–308.
- Sinha, S.R.; Kossoff, E.H. The ketogenic diet. Neurology 2005, 11, 161–170.
- Wheless, J.W. History of the ketogenic diet. Epilepsia 2008, 49 (Suppl. S8), 3–5.
- Lima, P.A.; Sampaio, L.P.; Damasceno, N.R. Neurobiochemical mechanisms of a ketogenic diet in refractory epilepsy. Clinics 2014, 69, 699–705.
- Allen, B.G.; Bhatia, S.K.; Anderson, C.M.; Eichenberger-Gilmore, J.M.; Sibenaller, Z.A.; Mapuskar, K.A.; Schoenfeld, J.D.; Buatti, J.M.; Spitz, D.R.; Fath, F.A. Ketogenic diets as an adjuvant cancer therapy: History and potential mechanism. Redox Biol. 2014, 2, 963–970.
- Kossoff, E.H.; Zupec-Kania, B.A.; Auvin, S.; Ballaban-Gil, K.R.; Christina Bergqvist, A.G.; Blackford, R.; Buchhalter, J.R.; Caraballo, R.H.; Cross, J.H.; Dahlin, M.G.; et al. Optimal clinical management of children receiving dietary therapies for epilepsy: Updated recommendations of the International Ketogenic Diet Study Group. Epilepsia Open 2018, 3, 175–192.
- Ferreira, L.; Lisenko, K.; Barros, B.; Zangeronimo, M.; Pereira, L.; Sousa, R. Influence of medium-chain triglycerides on consumption and weight gain in rats: A systematic review. J. Anim. Physiol. Anim. Nutr. 2014, 98, 1–8.
- Giordano, C.; Marchiò, M.; Timofeeva, E.; Biagini, G. Neuroactive peptides as putative mediators of antiepileptic ketogenic diets. Front. Neurol. 2014, 5, 63.
- Kossoff, E.H.; Dorward, J.L. The modified Atkins diet. Epilepsia 2008, 49 (Suppl. S8), 37–41.
- Miranda, M.J.; Turner, Z.; Magrath, G. Alternative diets to the classical ketogenic diet—Can we be more liberal? Epilepsy Res. 2012, 100, 278–285.
- Rui, L. Energy metabolism in the liver. Compr. Physiol. 2014, 4, 177.
- Robinson, A.M.; Williamson, D.H. Physiological roles of ketone bodies as substrates and signals in mammalian tissues. Physiol. Rev. 1980, 60, 143–187.
- Leonard, T. The physiology of ketosis and the ketogenic diet. South. Afr. J. Anaesth. Analg. 2020, 26, S94–S97.
- Laffel, L. Ketone bodies: A review of physiology, pathophysiology and application of monitoring to diabetes. Diabetes/Metab. Res. Rev. 1999, 15, 412–426.
- McGarry, J.D.; Woeltje, K.F.; Kuwajima, M.; Foster, D.W. Regulation of ketogenesis and the renaissance of carnitine palmitoyltransferase. Diabetes/Metab. Rev. 1989, 5, 271–284.
- Arnold, A.; Ali, A.; Kaka, N.; Kakodkar, P. Ketogenic Diet: Biochemistry, Weight Loss and Clinical Applications. Nutri. Food Sci. Int. J. 2020, 10, 555782.
- Krebs, H. The regulation of the release of ketone bodies by the liver. Adv. Enzym. Regul. 1966, 4, 339–353.
- Bilsborough, S.A.; Crowe, T. Low carbohydrate diets: What are the potential short and long term health implications? Asia Pac. J. Clin. Nutr. 2003, 12, 397–404.
- Adam-Perrot, A.; Clifton, P.; Brouns, F. Low-carbohydrate diets: Nutritional and physiological aspects. Obes. Rev. Off. J. Int. Assoc. Study Obes. 2006, 7, 49–58.
- Laeger, T.; Metges, C.C.; Kuhla, B. Role of β-hydroxybutyric acid in the central regulation of energy balance. Appetite 2010, 54, 450–455.
- Glew, H.R. You can get there from here: Acetone, anionic ketones and even-carbon fatty acids can provide substrates for gluconeogenesis. Niger. J. Physiol. Sci. 2010, 25, 2–4.
- McGarry, J.; Foster, D. Regulation of hepatic fatty acid oxidation and ketone body production. Annu. Rev. Biochem. 1980, 49, 395–420.
- Masino, S.A.; Rho, J.M. Mechanisms of ketogenic diet action. In Jasper’s Basic Mechanisms of the Epilepsies , 4th ed.; National Center for Biotechnology Information (US): Bethesda, MD, USA, 2012.
- Grabacka, M.; Pierzchalska, M.; Dean, M.; Reiss, K. Regulation of ketone body metabolism and the role of PPARα. Int. J. Mol. Sci. 2016, 17, 2093.