Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Protein kinase CK2 (CK2) is a ubiquitous holoenzyme involved in a wide array of developmental processes. The involvement of CK2 in events such as neurogenesis, cardiogenesis, skeletogenesis, and spermatogenesis is essential for the viability of almost all organisms, and its role has been conserved throughout evolution.
- casein kinase II
- development
- cancer
1. CK2 and Development
CK2 is highly expressed and prominent throughout many stages of development. During early embryogenesis, CK2 activity was identified first at embryonic day 11 (E11) in mice, rats, and chickens [1][2][3]. Continuously after this day, CK2 regulates the formation of various organ systems. Specifically, during neurogenesis and regarding the negative regulation of neuron differentiation, CK2 phosphorylates Groucho/Transducin-like enhancer (TLE1) to limit the formation of neurons [4]. CK2α null mice display improper neural tube formation, emphasizing the essential role of CK2 in this process [5]. Furthermore, CK2 contributes to memory formation, and the overexpression of this kinase is correlated with improved long-term memory [6]. In cardiogenesis, CK2, specifically CK2α’, is essential for the proper formation of the heart, and knockouts of this subunit lead to irregular cardiogenesis, as well as death in mid-gestation [5]. Similarly, CK2 is critical for proper spermatogenesis and the fertility of organisms [7]. While CK2α’−/− mice remain viable, male mice are infertile and incapable of producing offspring, as germ cells do not form properly [5][8][9]. Moreover, CK2α’ is highly expressed in testis, and a lack of CK2α’ also leads to oligozoospermia [8]. CK2 is also required for the formation of the skeleton during development, specifically at E17.5 [10]. It has also been demonstrated that the conditional deletion of the gene encoding beta subunit of CK2 (Csnk2b) leads to the shortening of the limbs, improper endochondral ossification, and lethality in mice [10]. Therefore, CK2 is critical for many stages of development and the production of viable organisms (Figure 1).
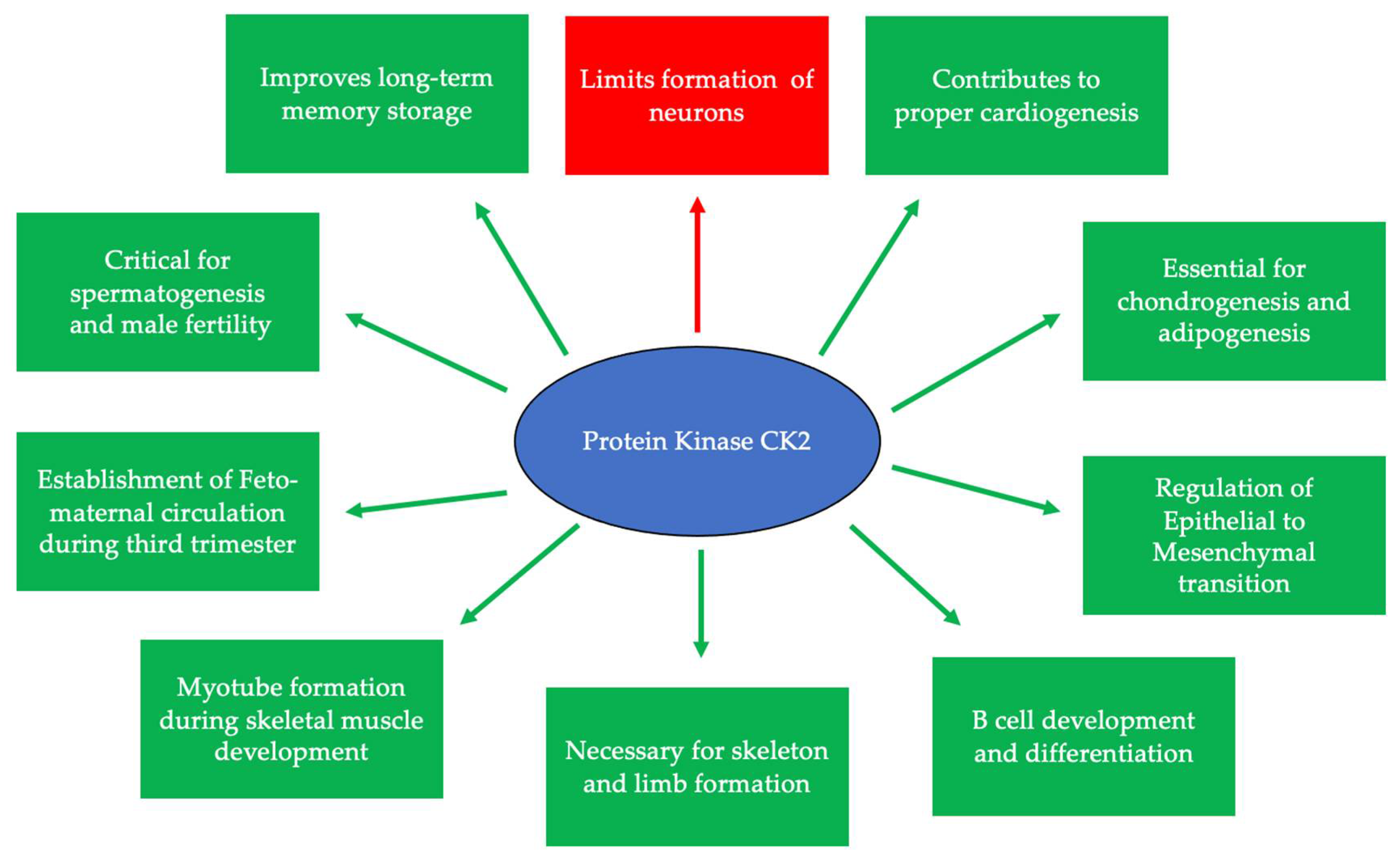
Figure 1. CK2 is expressed at embryonic day 11 and is critical for many developmental processes. Specifically, CK2 is necessary for limiting neurogenesis and preventing the excessive differentiation of neurons. In addition, CK2 expression promotes long-term memory storage. Further, this protein is essential for skeletogenesis, chondrogenesis, adipogenesis, and proper limb formation. CK2 contributes to spermatogenesis, and the inhibition of its expression leads to infertility. Finally, CK2 is important for B cell differentiation and development, myotube formation, the regulation of the epithelial-to-mesenchymal transition (EMT), and the establishment of circulation between the fetus and the mother during the third trimester.
There are three classifications of substrates for CK2. Class I substrates are equally phosphorylated by the holoenzyme and free catalytic subunits, whereas Class II substrates are phosphorylated by only free catalytic subunits but not by the holoenzyme. Further, substrates of Class III are phosphorylated preferentially by the holoenzyme but not by free catalytic subunits. Thus, the phosphorylation of these different classes varies during development (Figure 2) [11]. To study the role of the individual subunits during development, specific subunit knockouts were induced.
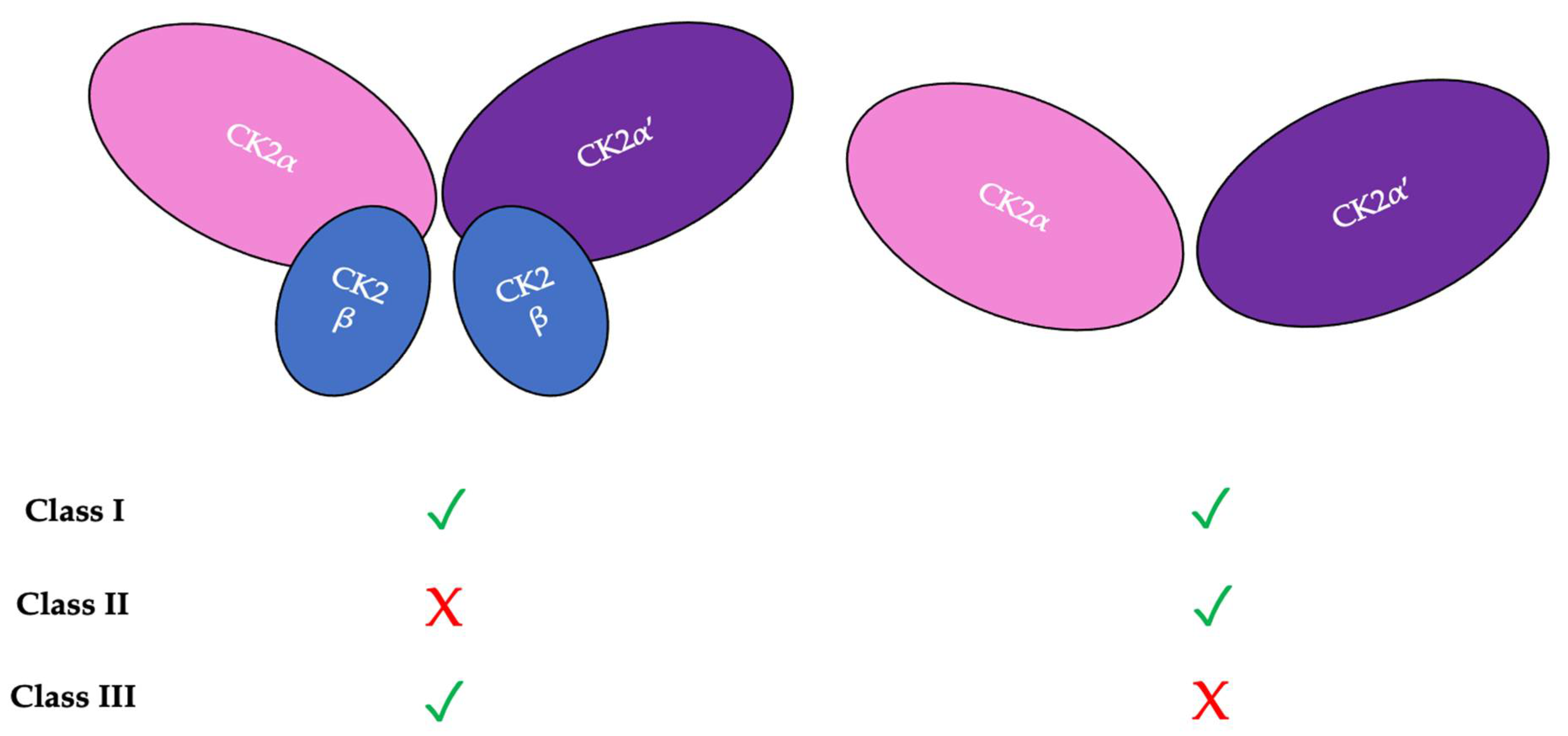
Figure 2. There are three classifications of CK2 substrates. Class I substrates are identified as proteins that are equally phosphorylated by the holoenzyme and individually by the catalytic subunits. Class II substrates are specifically phosphorylated by the catalytic submits of CK2 but not by the holoenzyme. Class III substrates are preferentially targeted and phosphorylated by the holoenzyme but not by the catalytic subunits of CK2.
2. CK2 Expression and the Progression of Diseases
The ubiquitous expression of CK2 occurs within almost all cellular compartments [12]. The continued identification of its newer substrates broadens the understanding of the processes in which this protein is involved [13]. Alternative forms of this holoenzyme, such as single catalytic or regulatory subunits, have differential expression and functions within tissues. The CK2α’ subunit has enhanced mRNA expression in the testis, skin, and across brain tissue. Further, the CKα and CK2β subunits have a more ubiquitous expression within tissues [14]. Oligomeric super-structures of the CK2 holoenzyme also exist, in which this protein controls the exposure of the catalytic active sites and subsequent enzymatic activity [15]. The balance between the number of catalytic CK2α and regulatory CK2β subunits is maintained by a mechanism whereby CK2α activates the transcription factors for CK2β when its levels are depleted. Once sufficient levels of CK2β are obtained, transcription is inhibited by holoenzyme formation, which then disables the interaction between CK2α and its own transcription factors [16].
2.1. Dynamic Localization of CK2
The subcellular localization of CK2 determines its regulation, which predominantly includes the cytoplasm, plasma membrane, and nucleus. Interactor proteins and substrates of CK2 are located predominantly in these three cellular compartments [17]. The localization of CK2 varies during cell cycle progression and is sensitive to external stimuli, such as radiation, hypoxia, and stress. Exposure to ionizing radiation (IR) was identified to alter CK2α subunit localization from the cytoplasm to the nucleus in human non-small cell lung cancer cell lines, including H460, A549, and PC9 (Figure 3). The nuclear localization is reported to be transient. In addition, there is an increase in the kinase activity of CK2 during the S phase when cells are exposed to IR. Furthermore, with the pharmacological inhibition of CK2, there is an increase in IR sensitivity [18][19]. This emphasizes the role of CK2 in transmitting radiation stimuli to the nucleus and its involvement in the DNA damage response. Similarly, in M059K human glioma cells, it colocalizes with the IR-induced DNA damage sites [20]. Resistance to IR treatment could be attributed to the dynamic localization of this kinase. During hypoxia, CK2 activity is seen to increase without an increase in its expression, but rather by the transport of catalytic α and α’ subunits to the nucleus (Figure 4).
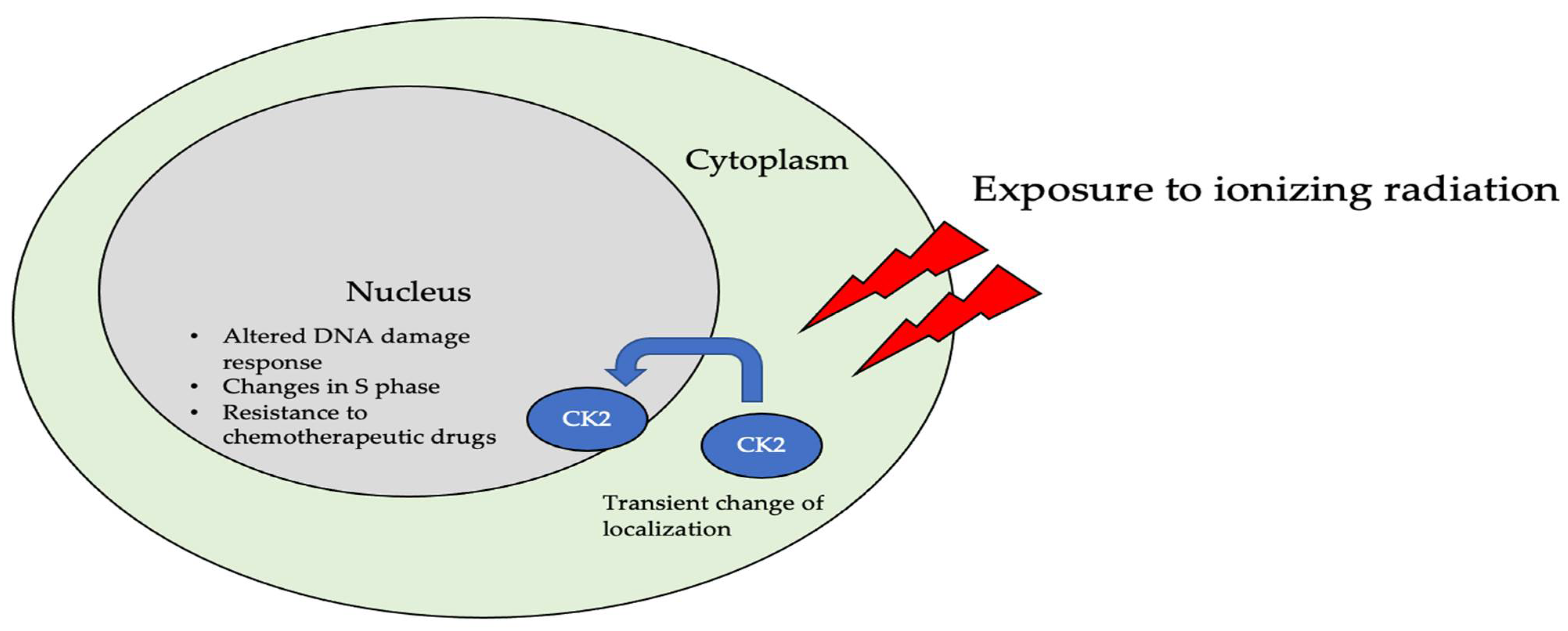
Figure 3. The localization of CK2 changes when cells are exposed to ionizing radiation (IR). Specifically, CK2α is translocated from the cytoplasm to the nucleus in multiple cell lines, including A549, H460, PC9, and M059K. Further, upon radiation exposure, the overall kinase activity of CK2 increases, emphasizing its role in the DNA-damage-repair response, as it colocalizes with DNA in the nucleus of cells.
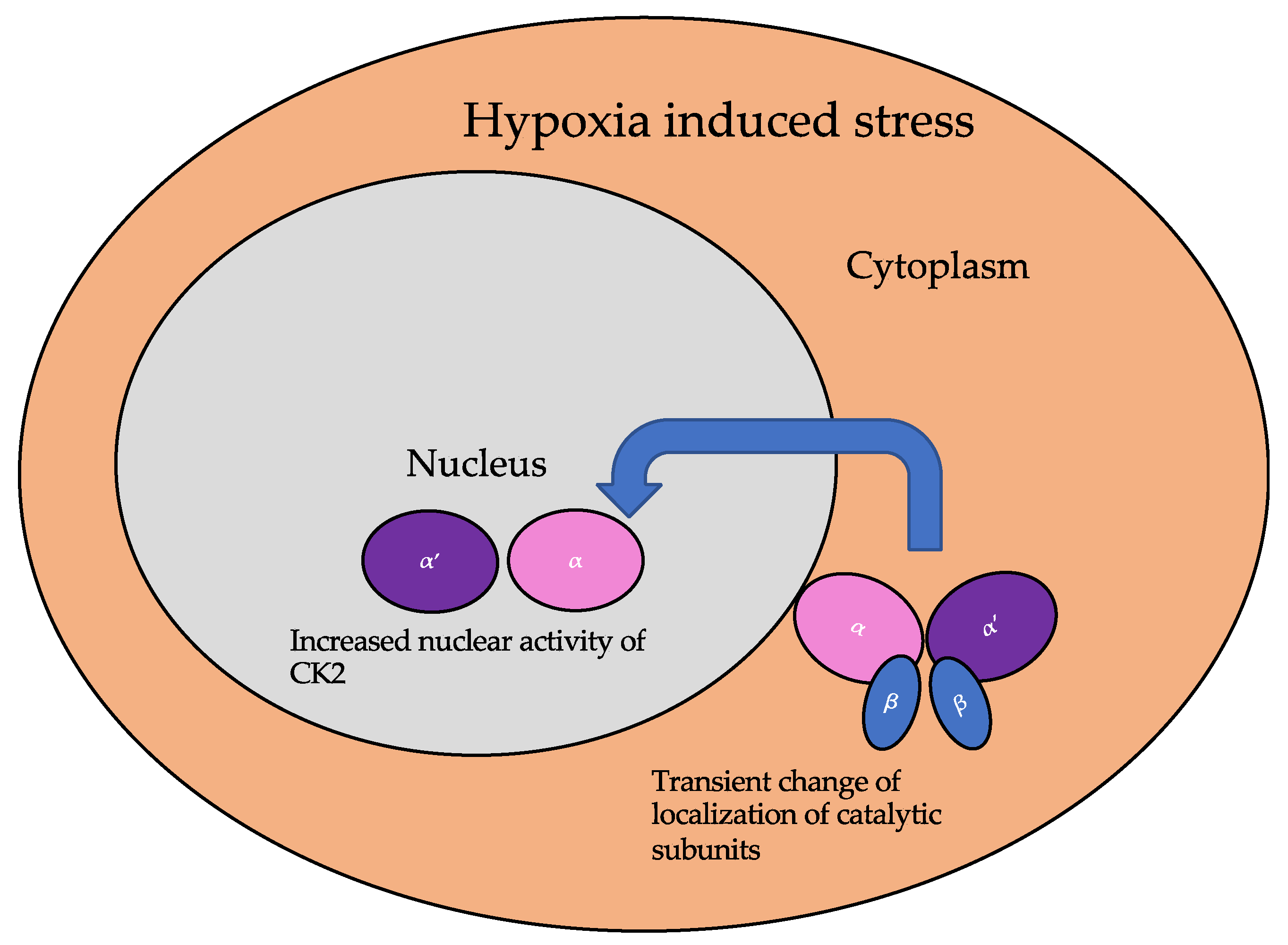
Figure 4. The localization of the catalytic subunits of CK2 is altered during hypoxia. Specifically, these subunits are translocated to the nucleus transiently to aid the cellular response to hypoxia.
2.2. CK2 in Diseases
CK2 is associated with the etiology of many diseases due to its pleiotropic nature and involvement in almost all cellular process. The diseases associated with the aberrant expression or function of this kinase include cancer, multiple sclerosis, cystic fibrosis, diabetes mellitus, neurological disorders, cardiac hypertrophy, and inflammation [21]. The examples delineated below describe the involvement of CK2 in disease progression based on unique mechanisms specific for the disease. The implication of aberrant CK2 expression and/or function is apparent in several studies.
2.2.1. CK2 in Cancer Progression
CK2 regulates the progression of at least 15 cancer-related proteins including the tumor suppressor p53, histone-modification enzymes such as HDAC1 and HDAC2, and NF-κB subunit RelA [22]. The interaction of additional direct interactors and indirectly regulated proteins remains unclear. There is an upregulation of CK2 during the tumor progression of many cancers and leukemia [23][24]. CK2 itself can be used as a prognostic marker for some cancer types[25]. Proteomic analyses of cell lines indicates that the catalytic CK2α subunit is predominantly expressed in several cancerous cell lines, including U2OS from human osteosarcoma, epidermoid squamous cell carcinoma, brain glioblastoma A549 from lung carcinoma, GAMG from glioblastoma, HEK293 from embryonic kidney cells, HeLa from cervical carcinoma, HepG2 from hepatoma, Jurkat from
As CK2 regulates the activation of several proteins required for the cell cycle, the consequences of its overexpression are exacerbated in solid tumors and leukemias [26]. CK2 overexpression and/or overactivity results in the abnormal phosphorylation of its targets. One example is tumor suppressor Ikaros, which is a transcriptional repressor of the genes encoding for Bcl-2-like protein 1 (BCL2L1, PIK3CD, and PIKFYVE) [27][28].
The central role of CK2 in pathway networks for cell cycle regulation and proliferation makes this protein an interesting pharmacological target for treatment of cancer.
2.2.2. CK2 and Diabetes and Obesity
In pancreatic β cells, a G-protein coupled-receptor muscarinic 3 receptor (M3R) regulates insulin release and homeostasis. CK2 is a negative modulator of M3R. Its phosphorylation of the M3Rs on β-cells inhibits insulin release [29]. Therefore, by phosphorylating the M3Rs, CK2 prevents the release of insulin. However, the abnormal activity of CK2 is linked to diabetes due to its role with these receptors.
Sirtuin1 (SIRT1) deacetylase is a substrate of CK2 and is a regulator for hepatic lipid metabolism. This protein modulates transcription as an energy sensor molecule with its deacetylase function. In diet-induced obesity, CK2 is highly overexpressed and therefore increases the activity of SIRT1. Aberrant phosphorylation of SIRT1 at serine 164 by CK2 inhibits the entry of the former into the nucleus and mildly impairs deacetylase function, thus hampering its regulatory function in hepatocytes, and has been linked to the onset of obesity [30].
2.2.3. CK2 and Heterotrophic Bone Formation
Heterotrophic bone formation in soft tissue due to injury, medication, or as a symptom of another disease can be a painful condition. CK2 phosphorylation stabilizes runt-related transcription factor 2 (RUNX2) by deubiquitylation through the CK2/herpesvirus-associated ubiquitin-specific protease (HAUSP)/RUNX2 pathway. This regulation is important during skeletal development for the fine-tuning of RUNX2, which is a master transcription factor during osteoblastogenesis. However, the overexpression or overactivity of CK2 during heterotrophic bone formation causes ectopic osteoblastogenesis from skeletal stem cells by the same mechanism for RUNX2 stabilization [10].
2.2.4. CK2 and Cardiovascular Diseases
Within the cardiovascular system, CK2α interacts with p27 to prevent its ubiquitination or degradation. Here, the stabilization of p27, which is an inhibitor of cell progression, leads to its accumulation in cytoplasm and the subsequent apoptosis of the cell. Further, CK2α is responsive to external growth stimuli to regulate its interaction with p27. The interaction between CK2α and p27 is interrupted during cardiac hypertrophy [31]. Here, transcriptional reprogramming leading to fetal gene expression causes cardiac hypertrophy. In addition, this reprogramming is related to HDAC2 activity. Specifically, CK2α phosphorylates HDAC2 at serine 394, causing its activation, and leads to cardiac hypertrophy [32].
Patients with cardiac desynchrony are associated with higher risk for morbidity and mortality from heart failure compared to patients with synchronic heart function. Cardiac resynchronization therapy (CRT, pacemaker) is the only known non-pharmaceutical intervention that has long-term effects on this disorder. However, some patients do not respond to this intervention. To study molecular mechanisms for dyssynchronous heart failure (HFdys) and the impact of CRT, proteomic analysis was performed. The comparison of phosphoproteome of HFdys before and after CRT led to the discovery that CK2 signaling is activated during HFdys and that CRT reverses it [33]. The contribution of CK2 signaling in cardiac desynchrony is still an ongoing area of research.
2.2.5. CK2 and Neurodegenerative Disorders
The expression and activity of CK2 in brain cells is involved in neurodegenerative disorders [34]. Here, the overexpression of CK2 is associated with neurodegeneration [35]. Further, there are numerous CK2 substrates associated with brain development and function. In Alzheimer’s disease, CK2 colocalizes with the neurofibrillary tangles. The hyperphosphorylation of the SET protein at serine 9 leads to its improper localization in the cytoplasm caused by the activation of CK2 by tau or β-amyloid (Aβ). The overexpression of CK2 also causes cognitive deficits by impairing synaptic plasticity and synaptogenesis [36]. In Parkinson’s disease (PD), CK2 is localized to Lewy bodies and phosphorylates α-synuclein and synphilin-1 [37]. The treatment of PD using levodopa (L-DOPA) is known to induce L-DOPA-induced dyskinesis (LID). CK2 is important for the regulation of pathways leading to LID. Specifically, CK2α downregulation reduces the severity of LID [34].
Regarding dopaminergic signaling, CK2 is a negative regulator of the D1 receptor (D1R). It binds directly to the Gαs or Gαolf (Golf) subunits, and its knockout leads to the elevated expression of D1R on the plasma membrane. This leads to an increased response to D1 agonists [38]. With the CK2 conditional knockouts in medium spiny neurons, which express dopaminergic receptors, CK2α regulates the D1 signaling pathway in vivo [39].
2.2.6. CK2 and Neurological Disorders
Further, in the neuropsychiatric disorder Tourette syndrome (TS), a mutation in the SLITRK1 gene is present. The gene product is a membrane-bound protein that regulates synapse formation. The symptoms of TS arise early in the childhood, and affected individuals often suffer from attention-deficit/hyperactivity disorder (ADHD) and obsessive–compulsive disorder (OCD). CK2 phosphorylates the SLITRK1 protein at its 14-3-3 binding site, and a mutation at this phosphorylation site inhibits neurite formation [40].
2.2.7. CK2 in Infectious Diseases
Viral proteins are substrates of CK2. The presence of host kinase phosphorylation sites within these proteins can be useful for the virus to identify host cell status [41]. In the proteome of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), a total of 25 phosphorylation sites have been identified. CK2 is one of the main kinases identified to regulate these sites. There is also activation of CK2 signaling observed during SARS-CoV-2 infection [41].
The hepatitis C virus (HCV) nonstructural protein 5A (NS5A) plays an important role in viral particle assembly. Structural studies of the NS5A and CK2 complex using time-resolved nuclear magnetic resonance (NMR) indicate that there are four possible CK2 phosphorylation sites in the NS5A D3 domain [42]. The expression of hepatitis B virus (HBV) is regulated by the Human La protein (hLa). The phosphorylation of hLa at serine 366 by CK2 activates the protein and increases HBV expression. The pharmacological inhibition of CK2 by tetrabromobenzimidazole (TBBz) or a knockdown its gene results in a reduction of HBV expression [43].
In prion diseases, the pathogenic form of the prion protein (Prp) interferes with cellular processes, such as fast axonal transport (FAT), which causes an onset of neurodegenerative disorders. The intracellular accumulation of cellular prion protein (PrPC) and full-length PrP (PrP-FL) induces neuronal toxicity, causing severe ataxia in mice, and further, FAT is often inhibited. CK2 inhibits FAT in a similar pattern as in PrP-FL-induced symptoms [44][45][46][47].
2.2.8. Regulation of Immune Response by CK2
The regulation of the immune cell response is targeted for the use of CK2 inhibitors. However, delineating its role during development in the immune response under disease progression has been a challenge. The involvement of CK2 is being investigated during this disease response in ongoing studies.
During Listeria monocytogenes (Lm) infection, the adaptive and innate immune responses are involved. In myeloid cell-specific conditional knockout of CK2α in mice, host resistance to Lm infection is significantly increased without affecting myeloid cell development. Myeloid cell recruitment is also seen to be negatively regulated by CK2α [48].
Further, the expression of CK2 is increased in the inflamed mucosa of ulcerative colitis (UC) patients. It is important for maintaining reciprocal balance between Th17 and Treg cells [49]. Interestingly, CK2 activity is reduced, along with an increase in reactive oxygen species (ROS) generation, during acute colitis. NADPH oxidase 1 (NOX1), which is an important regulator of mucosal immunity, is dysregulated during inflammatory bowel disease. Further, CK2 is a suppressor of NOX1 activity [50].
2.2.9. CK2 and Senescence
CK2 activity and expression is downregulated in older animals. Further, the pharmacological inhibition of CK2 induces cellular senescence [51][52]. With artificial downregulation of CK2 by the activity of histone trimethylases, such as histone-lysine N-methyltransferase SUV39H1(SUV39h1), its activity is upregulated in senescence and leads to an increase in tri-methylation at the 9th lysine residue of the histone H3 (H3K9me3) and senescence-associated heterochromatin formation (SAHF). An increase in SAHF leads to the suppression of genes associated with cell cycle progression, such as cyclin D1 [53].
Next, in CK2α knockdown cell lines (MCF-7 and HCT116 cells), the expression of histone demethylases [JmjC domain-containing histone demethylase (JMJD2/KDM4) and lysine-specific demethylase 1 (LSD1/KDM1a)] is downregulated at the translational level. The ectopic expression of these histone demethylases suppressed SAHF and senescence-associated β-galactosidase activity. The effects of the translational downregulation of LSD1 due to the reduction in CK2 activity are complex, since it has non-histone substrates such as p53.
3. Clinical Applications of CK2
The implication of CK2 in the progression of many diseases has led to its popularity as a therapeutic target. Specifically, researchers have attempted to prevent the kinase activity of CK2, thereby to prevent pathological activity. In cells, CK2 contains pocket domains for adenosine triphosphate (ATP)-binding [54]. The pocket domain is essential for the activation and subsequent enzymatic activity of CK2. Therefore, blocking this domain may deactivate CK2. One such inhibitor under investigation is CX-4945, known as silmitasertib in the clinic. CX-4945 is currently in clinical trials to treat cancer and is being tested as an oral administration to patients to limit the activity of CK2. Specifically, CX-4945 functions as an ATP-competitive inhibitor, leading to cell cycle arrest and apoptosis [55]. Here, CX-4945 inhibits the activity of both CK2α and CK2α’ by binding to the ATP-binding site of these subunits with a higher affinity than ATP [54][56][57][58]. This competitive binding leads to the decreased expression of CK2 and its subsequent signaling, such as Smad and PI3K/Akt pathways [55][58] (Figure 5A). This decreased activation of these pathways by CX-4945 has inhibited the progressions of several cancers, including gastric cancer, renal cancer, hematological cancer, cholangiocarcinoma, basal cell carcinoma, and medulla blastoma [56][59][60][61]. CX-4945 is a promising therapeutic agent and has displayed minimal side-effects, such as diarrhea, nausea, and anemia, suggesting its role as a safe treatment [61][62].
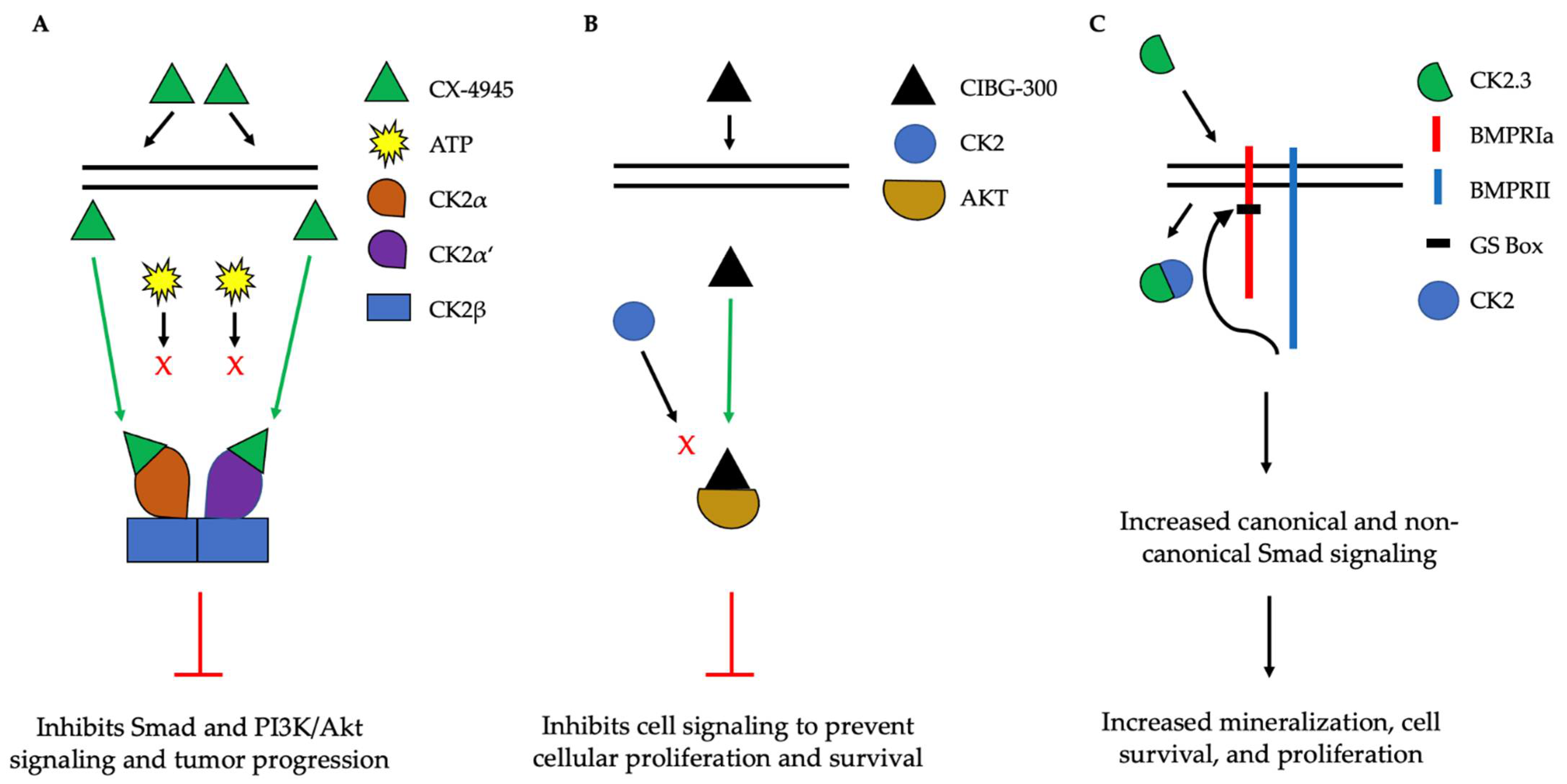
Figure 5. Simplified schematic demonstrating three therapeutics targeting the activity of CK2. (A). CX-4945 is a competitive ATP inhibitor that binds preferentially to ATP-pocket domains. Here, it binds to both CK2α and CK2α’, thereby preventing ATP from binding and activating CK2. CX-4945 has prevented tumor progression by inhibiting signaling pathways such as Smad and PI3K/AKT. (B). CIBG-300 functions by binding to conserved phosphorylation sequences on the substrates of CK2, such as Akt. This association prevents the phosphorylation of these substrates by CK2, leading to decreased cell signaling, proliferation, and survival. (C). CK2.3 is uptaken by cells and binds to CK2, therefore preventing its association with BMPRIa. As a result, BMPRII can phosphorylate BMPRIa, leading to the downstream activation of signaling pathways that induce mineralization and cell survival.
Another CK2 inhibitor, known as CIBG-300, is also currently in clinical trials as a therapeutic to treat cancer [63][64]. This inhibitor binds specifically to CK2 to prevent cellular proliferation; leads to decreased cell adhesion and migration; and promotes the apoptosis of the cells, specifically in cervical cancer, breast cancer, and colorectal cancer, both in vivo and in vitro [64][65][66][67][68][69][70][71]. Here, CIBG-300 functions by binding to the conserved phosphorylation sequences of CK2 substrates [68][72]. Subsequently, CK2 is unable to phosphorylate its downstream target; CIBG-300 thereby prevents the activity of various proteins involved in cancer progression and inhibits angiogenesis [68][69][70][73][74]. More interestingly, CIBG-300 inhibits the activation of Akt, PI3K, PTEN, and NF-κB signaling pathways, therefore decreasing cell proliferation and survival [75][76][77] (Figure 5B). Further, CIBG-300 treatment leads to limited side-effects including edema, hot flashes, tachycardia, lower abdominal pain, and bleeding [74]. Thus, this potential therapeutic has been efficient in clinical trials and holds promise to treat a wide array of cancers.
This entry is adapted from the peer-reviewed paper 10.3390/jdb10030031
References
- Schneider, H.R.; Reichert, G.H.; Issinger, O.G. Enhanced casein kinase II activity during mouse embryogenesis. Identification of a 110-kDa phosphoprotein as the major phosphorylation product in mouse embryos and Krebs II mouse ascites tumor cells. Eur. J. Biochem. 1986, 161, 733–738.
- Perez, M.; Grande, J.; Itarte, E. Developmental changes in rat hepatic casein kinases 1 and 2. Eur. J. Biochem. 1987, 170, 493–498.
- Maridor, G.; Park, W.; Krek, W.; Nigg, E.A. Casein kinase II. cDNA sequences, developmental expression, and tissue distribution of mRNAs for alpha, alpha’, and beta subunits of the chicken enzyme. J. Biol. Chem. 1991, 266, 2362–2368.
- Nuthall, H.N.; Joachim, K.; Stifani, S. Phosphorylation of serine 239 of Groucho/TLE1 by protein kinase CK2 is important for inhibition of neuronal differentiation. Mol. Cell Biol. 2004, 24, 8395–8407.
- Lou, D.Y.; Dominguez, I.; Toselli, P.; Landesman-Bollag, E.; O’Brien, C.; Seldin, D.C. The alpha catalytic subunit of protein kinase CK2 is required for mouse embryonic development. Mol. Cell Biol. 2008, 28, 131–139.
- Chao, C.C.; Ma, Y.L.; Lee, E.H. Protein kinase CK2 impairs spatial memory formation through differential cross talk with PI-3 kinase signaling: Activation of Akt and inactivation of SGK1. J. Neurosci. 2007, 27, 6243–6248.
- Mannowetz, N.; Kartarius, S.; Wennemuth, G.; Montenarh, M. Protein kinase CK2 and new binding partners during spermatogenesis. Cell Mol. Life Sci. 2010, 67, 3905–3913.
- Escalier, D.; Silvius, D.; Xu, X. Spermatogenesis of mice lacking CK2alpha’: Failure of germ cell survival and characteristic modifications of the spermatid nucleus. Mol. Reprod. Dev. 2003, 66, 190–201.
- Schwind, L.; Wilhelm, N.; Kartarius, S.; Montenarh, M.; Gorjup, E.; Götz, C. Protein kinase CK2 is necessary for the adipogenic differentiation of human mesenchymal stem cells. Biochim. Biophys. Acta 2015, 1853, 2207–2216.
- Kim, J.M.; Yang, Y.S.; Park, K.H.; Ge, X.; Xu, R.; Li, N.; Song, M.; Chun, H.; Bok, S.; Charles, J.F.; et al. A RUNX2 stabilization pathway mediates physiologic and pathologic bone formation. Nat. Commun. 2020, 11, 2289.
- Pinna, L.A. Protein kinase CK2: A challenge to canons. J. Cell Sci. 2002, 115, 3873–3878.
- Faust, M.; Montenarh, M. Subcellular localization of protein kinase CK2. A key to its function? Cell Tissue Res. 2000, 301, 329–340.
- Götz, C.; Montenarh, M. Protein kinase CK2 in development and differentiation. Biomed. Rep. 2017, 6, 127–133.
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Proteomics. Tissue-based map of the human proteome. Science 2015, 347, 1260419.
- Roffey, S.E.; Litchfield, D.W. CK2 Regulation: Perspectives in 2021. Biomedicines 2021, 9, 1361.
- Robitzki, A.; Bodenbach, L.; Voss, H.; Pyerin, W. Human casein kinase II. The subunit alpha protein activates transcription of the subunit beta gene. J. Biol. Chem. 1993, 268, 5694–5702.
- Filhol, O.; Cochet, C. Protein kinase CK2 in health and disease: Cellular functions of protein kinase CK2: A dynamic affair. Cell. Mol. Life Sci. 2009, 66, 1830–1839.
- Li, S.; Chen, P.; Yang, Q. Denosumab versus zoledronic acid in cases of surgically unsalvageable giant cell tumor of bone: A randomized clinical trial. J. Bone Oncol. 2019, 15, 100217.
- Li, Q.; Li, K.; Zhang, S.; Zhou, Y.; Hong, J.; Zhou, X.; Li, Z.; Wu, B.; Wu, G.; Meng, R. The effect of ionizing radiation on the subcellular localization and kinase activity of protein kinase CK2 in human non-small cell lung cancer cells. Int. J. Radiat. Biol. 2019, 95, 1462–1471.
- Olsen, B.B.; Wang, S.Y.; Svenstrup, T.H.; Chen, B.P.; Guerra, B. Protein kinase CK2 localizes to sites of DNA double-strand break regulating the cellular response to DNA damage. BMC Mol. Biol. 2012, 13, 7.
- Silva-Pavez, E.; Tapia, J.C. Protein Kinase CK2 in Cancer Energetics. Front. Oncol. 2020, 10, 893.
- Nuñez de Villavicencio-Diaz, T.; Rabalski, A.J.; Litchfield, D.W. Protein Kinase CK2: Intricate Relationships within Regulatory Cellular Networks. Pharmaceuticals 2017, 10, 27.
- Daya-Makin, M.; Sanghera, J.S.; Mogentale, T.L.; Lipp, M.; Parchomchuk, J.; Hogg, J.C.; Pelech, S.L. Activation of a tumor-associated protein kinase (p40TAK) and casein kinase 2 in human squamous cell carcinomas and adenocarcinomas of the lung. Cancer Res. 1994, 54, 2262–2268.
- Faust, R.A.; Gapany, M.; Tristani, P.; Davis, A.; Adams, G.L.; Ahmed, K. Elevated protein kinase CK2 activity in chromatin of head and neck tumors: Association with malignant transformation. Cancer Lett. 1996, 101, 31–35.
- Ahmad, K.A.; Wang, G.; Unger, G.; Slaton, J.; Ahmed, K. Protein kinase CK2—A key suppressor of apoptosis. Adv. Enzym. Regul. 2008, 48, 179–187.
- Mandato, E.; Manni, S.; Zaffino, F.; Semenzato, G.; Piazza, F. Targeting CK2-driven non-oncogene addiction in B-cell tumors. Oncogene 2016, 35, 6045–6052.
- Dovat, E.; Song, C.; Hu, T.; Rahman, M.A.; Dhanyamraju, P.K.; Klink, M.; Bogush, D.; Soliman, M.; Kane, S.; McGrath, M.; et al. Transcriptional Regulation of PIK3CD and PIKFYVE in T-Cell Acute Lymphoblastic Leukemia by IKAROS and Protein Kinase CK2. Int. J. Mol. Sci. 2021, 22, 819.
- Song, C.; Ge, Z.; Ding, Y.; Tan, B.H.; Desai, D.; Gowda, K.; Amin, S.; Gowda, R.; Robertson, G.P.; Yue, F.; et al. IKAROS and CK2 regulate expression of BCL-XL and chemosensitivity in high-risk B-cell acute lymphoblastic leukemia. Blood 2020, 136, 1520–1534.
- Rossi, M.; Ruiz de Azua, I.; Barella, L.F.; Sakamoto, W.; Zhu, L.; Cui, Y.; Lu, H.; Rebholz, H.; Matschinsky, F.M.; Doliba, N.M.; et al. CK2 acts as a potent negative regulator of receptor-mediated insulin release in vitro and in vivo. Proc. Natl. Acad. Sci. USA 2015, 112, E6818–E6824.
- Choi, S.E.; Kwon, S.; Seok, S.; Xiao, Z.; Lee, K.W.; Kang, Y.; Li, X.; Shinoda, K.; Kajimura, S.; Kemper, B.; et al. Obesity-Linked Phosphorylation of SIRT1 by Casein Kinase 2 Inhibits Its Nuclear Localization and Promotes Fatty Liver. Mol. Cell. Biol. 2017, 37, e00006-17.
- Hauck, L.; Harms, C.; An, J.; Rohne, J.; Gertz, K.; Dietz, R.; Endres, M.; von Harsdorf, R. Protein kinase CK2 links extracellular growth factor signaling with the control of p27(Kip1) stability in the heart. Nat. Med. 2008, 14, 315–324.
- Eom, G.H.; Cho, Y.K.; Ko, J.H.; Shin, S.; Choe, N.; Kim, Y.; Joung, H.; Kim, H.S.; Nam, K.I.; Kee, H.J.; et al. Casein kinase-2α1 induces hypertrophic response by phosphorylation of histone deacetylase 2 S394 and its activation in the heart. Circulation 2011, 123, 2392–2403.
- Stachowski, M.J.; Holewinski, R.J.; Grote, E.; Venkatraman, V.; Van Eyk, J.E.; Kirk, J.A. Phospho-Proteomic Analysis of Cardiac Dyssynchrony and Resynchronization Therapy. Proteomics 2018, 18, e1800079.
- Cortés, M.; Malave, L.; Castello, J.; Flajolet, M.; Cenci, M.A.; Friedman, E.; Rebholz, H. CK2 Oppositely Modulates l-DOPA-Induced Dyskinesia via Striatal Projection Neurons Expressing D1 or D2 Receptors. J. Neurosci. 2017, 37, 11930–11946.
- Perez, D.I.; Gil, C.; Martinez, A. Protein kinases CK1 and CK2 as new targets for neurodegenerative diseases. Med. Res. Rev. 2011, 31, 924–954.
- Zhang, Q.; Xia, Y.; Wang, Y.; Shentu, Y.; Zeng, K.; Mahaman, Y.A.R.; Huang, F.; Wu, M.; Ke, D.; Wang, Q.; et al. CK2 Phosphorylating I. Front. Mol. Neurosci. 2018, 11, 146.
- Ryu, M.Y.; Kim, D.W.; Arima, K.; Mouradian, M.M.; Kim, S.U.; Lee, G. Localization of CKII beta subunits in Lewy bodies of Parkinson’s disease. J. Neurol. Sci. 2008, 266, 9–12.
- Rebholz, H.; Nishi, A.; Liebscher, S.; Nairn, A.C.; Flajolet, M.; Greengard, P. CK2 negatively regulates Galphas signaling. Proc. Natl. Acad. Sci. USA 2009, 106, 14096–14101.
- Rebholz, H.; Zhou, M.; Nairn, A.C.; Greengard, P.; Flajolet, M. Selective knockout of the casein kinase 2 in d1 medium spiny neurons controls dopaminergic function. Biol. Psychiatry 2013, 74, 113–121.
- Kajiwara, Y.; Buxbaum, J.D.; Grice, D.E. SLITRK1 binds 14-3-3 and regulates neurite outgrowth in a phosphorylation-dependent manner. Biol. Psychiatry 2009, 66, 918–925.
- Bouhaddou, M.; Memon, D.; Meyer, B.; White, K.M.; Rezelj, V.V.; Correa Marrero, M.; Polacco, B.J.; Melnyk, J.E.; Ulferts, S.; Kaake, R.M.; et al. The Global Phosphorylation Landscape of SARS-CoV-2 Infection. Cell 2020, 182, 685–712.
- Secci, E.; Luchinat, E.; Banci, L. The Casein Kinase 2-Dependent Phosphorylation of NS5A Domain 3 from Hepatitis C Virus Followed by Time-Resolved NMR Spectroscopy. ChemBioChem 2016, 17, 328–333.
- Tang, J.; Zhang, Z.H.; Huang, M.; Heise, T.; Zhang, J.; Liu, G.L. Phosphorylation of human La protein at Ser366 by casein kinase II contributes to hepatitis B virus replication and expression in vitro. J. Viral Hepat. 2013, 20, 24–33.
- Wang, S.; Jones, K.A. CK2 controls the recruitment of Wnt regulators to target genes in vivo. Curr. Biol. 2006, 16, 2239–2244.
- Rane, N.S.; Yonkovich, J.L.; Hegde, R.S. Protection from cytosolic prion protein toxicity by modulation of protein translocation. EMBO J. 2004, 23, 4550–4559.
- Zamponi, E.; Buratti, F.; Cataldi, G.; Caicedo, H.H.; Song, Y.; Jungbauer, L.M.; LaDu, M.J.; Bisbal, M.; Lorenzo, A.; Ma, J.; et al. Prion protein inhibits fast axonal transport through a mechanism involving casein kinase 2. PLoS ONE 2017, 12, e0188340.
- Rane, N.S.; Kang, S.W.; Chakrabarti, O.; Feigenbaum, L.; Hegde, R.S. Reduced translocation of nascent prion protein during ER stress contributes to neurodegeneration. Dev. Cell 2008, 15, 359–370.
- Larson, S.R.; Bortell, N.; Illies, A.; Crisler, W.J.; Matsuda, J.L.; Lenz, L.L. Myeloid Cell CK2 Regulates Inflammation and Resistance to Bacterial Infection. Front. Immunol. 2020, 11, 590266.
- Dong, G.; Yang, Y.; Zhang, H.; Yu, W.; He, H.; Dai, F.; Ma, C.; Wang, Y.; Zhu, F.; Xiong, H.; et al. Protein Kinase CK2 Maintains Reciprocal Balance Between Th17 and Treg Cells in the Pathogenesis of UC. Inflamm. Bowel Dis. 2022, 28, 830–842.
- Liu, D.; Marie, J.C.; Pelletier, A.L.; Song, Z.; Ben-Khemis, M.; Boudiaf, K.; Pintard, C.; Leger, T.; Terrier, S.; Chevreux, G.; et al. Protein Kinase CK2 Acts as a Molecular Brake to Control NADPH Oxidase 1 Activation and Colon Inflammation. Cell. Mol. Gastroenterol. Hepatol. 2022, 13, 1073–1093.
- Park, J.H.; Lee, J.H.; Park, J.W.; Kim, D.Y.; Hahm, J.H.; Nam, H.G.; Bae, Y.S. Downregulation of protein kinase CK2 activity induces age-related biomarkers in C. elegans. Oncotarget 2017, 8, 36950–36963.
- Ryu, S.W.; Woo, J.H.; Kim, Y.H.; Lee, Y.S.; Park, J.W.; Bae, Y.S. Downregulation of protein kinase CKII is associated with cellular senescence. FEBS Lett. 2006, 580, 988–994.
- Park, J.W.; Kim, J.J.; Bae, Y.S. CK2 downregulation induces senescence-associated heterochromatic foci formation through activating SUV39h1 and inactivating G9a. Biochem. Biophys. Res. Commun. 2018, 505, 67–73.
- Ferguson, A.D.; Sheth, P.R.; Basso, A.D.; Paliwal, S.; Gray, K.; Fischmann, T.O.; Le, H.V. Structural basis of CX-4945 binding to human protein kinase CK2. FEBS Lett. 2011, 585, 104–110.
- Pierre, F.; Chua, P.C.; O’Brien, S.E.; Siddiqui-Jain, A.; Bourbon, P.; Haddach, M.; Michaux, J.; Nagasawa, J.; Schwaebe, M.K.; Stefan, E.; et al. Pre-clinical characterization of CX-4945, a potent and selective small molecule inhibitor of CK2 for the treatment of cancer. Mol. Cell. Biochem. 2011, 356, 37–43.
- Chon, H.J.; Bae, K.J.; Lee, Y.; Kim, J. The casein kinase 2 inhibitor, CX-4945, as an anti-cancer drug in treatment of human hematological malignancies. Front. Pharmacol. 2015, 6, 70.
- Lee, J.Y.; Yun, J.S.; Kim, W.K.; Chun, H.S.; Jin, H.; Cho, S.; Chang, J.H. Structural Basis for the Selective Inhibition of Cdc2-Like Kinases by CX-4945. BiomMed Res. Int. 2019, 2019, 6125068.
- Son, Y.H.; Moon, S.H.; Kim, J. The protein kinase 2 inhibitor CX-4945 regulates osteoclast and osteoblast differentiation in vitro. Mol. Cells 2013, 36, 417–423.
- Siddiqui-Jain, A.; Drygin, D.; Streiner, N.; Chua, P.; Pierre, F.; O’Brien, S.E.; Bliesath, J.; Omori, M.; Huser, N.; Ho, C.; et al. CX-4945, an orally bioavailable selective inhibitor of protein kinase CK2, inhibits prosurvival and angiogenic signaling and exhibits antitumor efficacy. Cancer Res. 2010, 70, 10288–10298.
- Richter, A.; Sender, S.; Lenz, A.; Schwarz, R.; Hinz, B.; Knuebel, G.; Sekora, A.; Murua Escobar, H.; Junghanss, C.; Roolf, C. Influence of Casein kinase II inhibitor CX-4945 on BCL6-mediated apoptotic signaling in B-ALL in vitro and in vivo. BMC Cancer 2020, 20, 184.
- Kim, H.M.; Jeong, I.; Kim, H.J.; Kang, S.K.; Kwon, W.S.; Kim, T.S.; Park, K.H.; Jung, M.; Soong, J.; Lin, S.C.; et al. Casein Kinase 2 Inhibitor, CX-4945, as a Potential Targeted Anticancer Agent in Gastric Cancer. Anticancer Res. 2018, 38, 6171–6180.
- Martins, L.R.; Lúcio, P.; Melão, A.; Antunes, I.; Cardoso, B.A.; Stansfield, R.; Bertilaccio, M.T.; Ghia, P.; Drygin, D.; Silva, M.G.; et al. Activity of the clinical-stage CK2-specific inhibitor CX-4945 against chronic lymphocytic leukemia. Leukemia 2014, 28, 179–182.
- Borgo, C.; D’Amore, C.; Sarno, S.; Salvi, M.; Ruzzene, M. Protein kinase CK2: A potential therapeutic target for diverse human diseases. Signal Transduct. Target. Ther. 2021, 6, 183.
- Perea, S.E.; Baladrón, I.; Valenzuela, C.; Perera, Y. CIGB-300: A peptide-based drug that impairs the Protein Kinase CK2-mediated phosphorylation. Semin. Oncol. 2018, 45, 58–67.
- Benavent Acero, F.; Capobianco, C.S.; Garona, J.; Cirigliano, S.M.; Perera, Y.; Urtreger, A.J.; Perea, S.E.; Alonso, D.F.; Farina, H.G. CIGB-300, an anti-CK2 peptide, inhibits angiogenesis, tumor cell invasion and metastasis in lung cancer models. Lung Cancer 2017, 107, 14–21.
- Gottardo, M.F.; Capobianco, C.S.; Sidabra, J.E.; Garona, J.; Perera, Y.; Perea, S.E.; Alonso, D.F.; Farina, H.G. Preclinical efficacy of CIGB-300, an anti-CK2 peptide, on breast cancer metastasic colonization. Sci. Rep. 2020, 10, 14689.
- Perea, S.E.; Reyes, O.; Baladron, I.; Perera, Y.; Farina, H.; Gil, J.; Rodriguez, A.; Bacardi, D.; Marcelo, J.L.; Cosme, K.; et al. CIGB-300, a novel proapoptotic peptide that impairs the CK2 phosphorylation and exhibits anticancer properties both in vitro and in vivo. Mol. Cell. Biochem. 2008, 316, 163–167.
- Perera, Y.; Ramos, Y.; Padrón, G.; Caballero, E.; Guirola, O.; Caligiuri, L.G.; Lorenzo, N.; Gottardo, F.; Farina, H.G.; Filhol, O.; et al. CIGB-300 anticancer peptide regulates the protein kinase CK2-dependent phosphoproteome. Mol. Cell. Biochem. 2020, 470, 63–75.
- Lian, H.; Su, M.; Zhu, Y.; Zhou, Y.; Soomro, S.H.; Fu, H. Protein Kinase CK2, a Potential Therapeutic Target in Carcinoma Management. Asian Pac. J. Cancer Prev. 2019, 20, 23–32.
- Farina, H.G.; Benavent Acero, F.; Perera, Y.; Rodríguez, A.; Perea, S.E.; Castro, B.A.; Gomez, R.; Alonso, D.F.; Gomez, D.E. CIGB-300, a proapoptotic peptide, inhibits angiogenesis in vitro and in vivo. Exp. Cell Res. 2011, 317, 1677–1688.
- Iegre, J.; Atkinson, E.L.; Brear, P.D.; Cooper, B.M.; Hyvönen, M.; Spring, D.R. Chemical probes targeting the kinase CK2: A journey outside the catalytic box. Org. Biomol. Chem. 2021, 19, 4380–4396.
- Perea, S.E.; Reyes, O.; Puchades, Y.; Mendoza, O.; Vispo, N.S.; Torrens, I.; Santos, A.; Silva, R.; Acevedo, B.; López, E.; et al. Antitumor effect of a novel proapoptotic peptide that impairs the phosphorylation by the protein kinase 2 (casein kinase 2). Cancer Res. 2004, 64, 7127–7129.
- Rodríguez-Ulloa, A.; Ramos, Y.; Gil, J.; Perera, Y.; Castellanos-Serra, L.; García, Y.; Betancourt, L.; Besada, V.; González, L.J.; Fernández-de-Cossio, J.; et al. Proteomic profile regulated by the anticancer peptide CIGB-300 in non-small cell lung cancer (NSCLC) cells. J. Proteome Res. 2010, 9, 5473–5483.
- Solares, A.M.; Santana, A.; Baladrón, I.; Valenzuela, C.; González, C.A.; Díaz, A.; Castillo, D.; Ramos, T.; Gómez, R.; Alonso, D.F.; et al. Safety and preliminary efficacy data of a novel casein kinase 2 (CK2) peptide inhibitor administered intralesionally at four dose levels in patients with cervical malignancies. BMC Cancer 2009, 9, 146.
- Husain, K.; Williamson, T.T.; Nelson, N.; Ghansah, T. Protein kinase 2 (CK2): A potential regulator of immune cell development and function in cancer. Immunol. Med. 2021, 44, 159–174.
- Martins, L.R.; Perera, Y.; Lúcio, P.; Silva, M.G.; Perea, S.E.; Barata, J.T. Targeting chronic lymphocytic leukemia using CIGB-300, a clinical-stage CK2-specific cell-permeable peptide inhibitor. Oncotarget 2014, 5, 258–263.
- Cirigliano, S.M.; Díaz Bessone, M.I.; Berardi, D.E.; Flumian, C.; Bal de Kier Joffé, E.D.; Perea, S.E.; Farina, H.G.; Todaro, L.B.; Urtreger, A.J. The synthetic peptide CIGB-300 modulates CK2-dependent signaling pathways affecting the survival and chemoresistance of non-small cell lung cancer cell lines. Cancer Cell Int. 2017, 17, 42.
This entry is offline, you can click here to edit this entry!