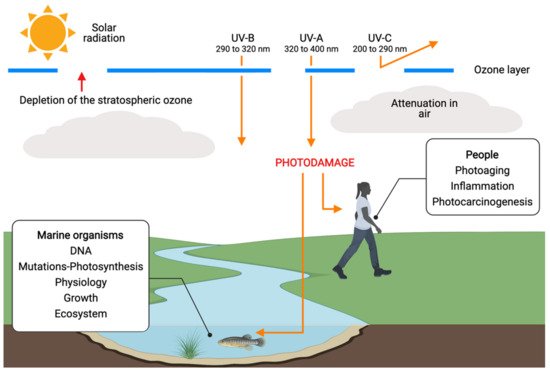
To prevent skin cancer and other issues such as sunburns, photoaging, hyperpigmentation, and skin wrinkles, the use of sunscreen is recommended. The UV-B radiation that reaches the Earth is mainly responsible for these types of skin damage [
10]. The most known and currently accepted method of sunscreen rating is the sun protection factor (SPF) system. However, not only the sun protection factor is a key parameter in measuring how effective a sunscreen is, but also the immune protection factor plays an important and very different role. This one correlates to the UV-A protective capability of the sunscreen for the prevention of UV immunosuppression [
6]. The inhibition of lipid peroxidation (LPO) species formed due to photooxidative damage is a good measure for UV protection [
11]. There are sunscreens that can decrease the LPO in the
stratum corneum, helping to establish the efficacy of anti-UV and antioxidant agents [
12].
2. UV-Resistant Microalgae
Different species of microalgae show a wide range of susceptibility to UV-B radiation [
20] and some have adapted to UV radiation over the years. Marine organisms, including eukaryotic microalgae and cyanobacteria, during evolution, have developed physiological and biochemical traits for defense and/or tolerance to UV rays [
26]. Although this is an exploitable feature of microalgae, limited information is available on the molecular mechanisms involved in stress response, adaptation, and resilience of microalgae to UV radiation. Some of the research made regarding this feature is mentioned in the following section. Núñez-Pons et al. [
27] reported some microalgae with UV resistant properties. For instance,
Eutreptiella sp. from the Southern Ocean, under variable light conditions, was successfully protected by the photoprotective pigments. This research also mentioned the Antarctic microalgae
Chaetoceros dichaeta,
Phaeocystis antarctica, and
Polarella glacialis, these were subjected to acclimation to photosynthetically active radiation (PAR) and showed resistance to UV-B rays. Among these Antarctic microalgae,
Polarella glacialis was shown to possess a very high xanthophyll to chlorophyll ratio [
27], which implies a high content of UV-absorbing compounds. Buma et al. support these findings stating that
P. antarctica and
P. glacialis showed strong linear induction of UV-absorbing compounds at increasing PAR intensities [
28].
So far, several species, e.g.,
Coelastrella rubescens,
Coelastrella terestris,
Enallax coelastroides,
Scotiella chlorelloidea,
Ankistrodesmus spiralis,
Chlorella minutissima,
Chlorella sorokiniana, and
Pseudococcomyxa sp. have been reported as rich sources of MAAs [
29]. As it was mentioned before, these act as effective UV-B screens. This report demonstrates the accumulations of MAAs when UV-B is induced. Kováčik et al. [
30] states the tolerance of Scenedesmus to UV-B as well as to UV-A and UV-C. The effects of UV rays on microalgae are investigated by studying the effect of UV stress on the fatty acid and lipid class composition in two marine microalgae
Pavlova lutheri, currently known as
Diacronema lutheri, and
Odontella aurita [
31]. The results obtained in the investigation for
Odontella aurita suggest that this species is more resistant and seems to be able to partially acclimate to UV rays based on the analysis of the lipids present in the microalga [
31]. This finding was supported in other research [
32] which mentions that the microalgae
Odontella aurita was able to acclimate to UV rays’ stress.
3. UV-Induced Microalgae Biosynthesis
3.1. Mycosporine-Like Amino Acids (MAAs)
MAAs are small intracellular, colorless water-soluble secondary metabolites of low molecular weight, commonly found in marine environments. These compounds have absorption maxima in the UV region between 310 and 360 nm, they can dissipate radiation as heat without producing ROS. Under fixed light, MAA biosynthesis seems the most effective photoprotective mechanism [
45]. More specifically, the biosynthesis of MAAs happens principally via the shikimate pathway [
51,
52,
53,
54,
55]. MAAs have been found in tropical to Antarctic waters and in a variety of organisms, spanning from cyanobacteria, microalgae, fungi, as well as macroalgae, and animals. In addition to protecting cells from mutation caused by UV rays and free radicals, they are also effective antioxidant molecules [
27].
More than thirty different MAAs have been identified from marine and terrestrial environments [
56]. The vital roles of different MAAs in protecting mechanisms by virtue of their UV-absorbing and antioxidant function have been reported in several microalgae [
26]. MAAs are considered the best photoprotective compound in microalgae due to their strong absorption capability in the UV spectrum, photostability in both fresh and saline water in the presence of photosensitizers, their antioxidant properties, and resistance to several abiotic stressors such as temperature, UV radiation, and various solvents and pH. For these reasons, they potentially can prevent UV-induced skin damage [
57]. It has been reported that MAAs protect against DNA damage, effectively block thymine dimer formation by UV radiation, protect the cells from UV-induced cell death, they also promote the proliferation of human skin fibroblast cells [
58]. These characteristics make MAAs an excellent alternative for green biotech applications in sunscreens.
3.2. Carotenoids
Carotenoids are structurally and functionally very diverse natural pigments and important components of the photosynthetic apparatus, they enhance cellular photosynthetic production and provide photooxidative protection. One of the most characteristic features of carotenoids is their strong coloration, and this characteristic is crucial for the proper functioning of light absorption in photosynthetic organisms and photoprotection in all living organisms. This feature plays an important role in photoprotection against UV-A radiation in cyanobacteria. Several carotenoids such as canthaxanthin, echinenone, myxoxanthophyll, and zeaxanthin with protective functions against photo-oxidative damage have been reported in several cyanobacteria and other green algae species, e.g.,
Coelastrella rubescens [
42]. Nearly all carotenoids absorb light in the 400–500 nm range. Their quenching properties allow the dissipation of excess energy from UV-B [
27]. Carotenoids play an important role in absorbing light energy for use in photosynthesis as well as protecting chlorophyll from being damaged by visible or UV light. The content of total carotenoids has been found to correlate with their protection against UV light [
25].
3.3. Sporopollenin
Sporopollenin is an acetolysis-resistant inert biopolymer possessing a complicated structure with aliphatic (mainly isoprenoid) and aromatic components variably present. It is found in the cell wall of some algae and in plant pollens and spores [
21]. It was reported that species of microalgae that were highly tolerant to UV radiation had substantial amounts of sporopollenin. This compound functions as an antimicrobial agent, confers rigidness to the cell wall, and may further protect from UV rays by increasing the optical density, whereas species containing little or no sporopollenin were highly UV radiation susceptible. In the pollen and spores of Antarctic plants, sporopollenin is considered a bio-indicator of solar UV-B and a valuable archive for the reconstruction of past solar UV-B [
20]. Sporopollenin-like substances seem to be the key protectors of some microalgae against UV. Due to high stability, sporopollenin-like compounds provide constant UV protection [
42].
3.4. Scytonemin
Scytonemin is a yellow-brown lipid soluble sheath pigment that absorbs maximally in the UV-A and UV-C regions, but with some absorbance in the UV-B region. It has an in vivo absorption maximum of 370 nm. It is synthesized by both temperature increase, and oxidative stress combined with UV-A [
27]. Other reports determine that scytonemin is suspected to be synthesized from metabolites of aromatic amino acid biosynthesis [
21]. Scytonemin is located in the extracellular polysaccharide sheath of some cyanobacteria acting as a passive sunscreen against UV radiation. It can also reduce about 90% of solar UV-A radiation that can reach the cell. Microalgal scytonemin appears restricted to cyanobacteria. The high stability of this compound against stressors such as temperature and UV-A radiation and its capability to undertake UV screening even after prolonged physiological inactivity such as desiccation makes it a strong candidate for use as a natural UV screening compound for humans [
21].
3.5. Phenolic Compounds
Phenolic compounds, which are also present in microalgae, also likely contribute to UV tolerance. Phenolic compounds in brown algae contribute considerably to protecting sensitive life stages from irradiation [
59]. These compounds are one of the largest groups of secondary metabolites present in plants, ranging from simple aromatic rings to more complex molecules and comprising flavonoids, phenolic acids, tannins, lignans, or coumarins. Phenolic compounds are also a part of the UV tolerance sector of interest, they are used in cosmetics as UV-protection and antioxidant agents [
60].
4. Bio-Carriers for Skin Applications
Controlled drug delivery is one of the most important areas of research in the creation of skin care products such as cosmeceuticals. It is desired for most drugs due to safety and efficacy reasons. By encapsulating such drugs into biodegradable polysaccharide materials, both improved bioavailability and controlled drug release is expected [
61]. Bio-based materials as bio-carriers are noted as key ingredients for the development of drug delivery systems [
62], while polysaccharides are the basis of the production of bio-based materials [
63]. However, due to their properties such as bioavailability, biodegradability, stability, availability, renewability, and low toxicity, polysaccharides can be part of pharmaceutical products, but they can also be applied as independent solid matrices in different forms (monoliths, beads, micro- or nanoparticles). Nanoparticles based on hydrophilic polymers are appropriate candidates for drug delivery [
64].
A good product formulation allows the user to obtain the greatest possible benefits from the product to be used. Different bio-carriers will perform different functions, some bio-carriers used in cosmetics that can be used for the formulation of sunscreen gels for their drug delivery potential are alginates, chitosan, gelatin, and cellulose to mention a few.
Table 2 shows a summary of some microalgae used along with bio-carriers for drug delivery [
65,
66,
67,
68,
69,
70,
71,
72,
73].
Table 2. Applications of bio-carriers with microalgae for drug delivery.
| Material Used |
Microalgae |
Component Loaded |
Formulation |
Purpose |
Types of Tests |
References |
| Chitosan |
Spirulina |
5-Fluorouracil |
Chitosan (1%(v/v)), Spirulina microalgae and genipin. |
Drug delivery |
In vitro cytotoxicity test |
[65] |
| Alginate and Chitosan |
Auxonochlorella
protothecoides |
Microalgae oil extract |
PVA solution (7–8% (w/v)) with sodium alginate (2% (w/v)) and PVA solution (7–8% (w/v)) with chitosan (2–3% (w/v)) |
Nanoparticle production to deliver bioactive compounds in microalgae |
In vitro release testing |
[66,67,68] |
| Alginate, PVA |
Botryococcus braunii and
Microcystis aeruginosa |
Microalgae oil extract |
PVA solution (8% (w/v)) and sodium alginate (2% (w/v)) |
Nanoparticle production to deliver bioactive compounds in microalgae |
In vitro release profile of nanoparticles |
[66,69] |
| Alginate, cellulose |
Chlorella vulgaris |
Tea tree essential oil |
Microfibrillated cellulose, nanofibrillated cellulose and carboxymethylcellulose (0.01% (m/v)) with alginate (2% (m/v)) |
Cosmetic application |
Differential scanning calorimetry |
[70] |
| Cellulose |
Chlorella pyrenoidosa |
- |
Microalgae powder, sodium dodecyl sulfate |
Sensor for pollutant detection |
- |
[71] |
| Gelatin |
Phaeodactylum tricornutum |
Microalgae extract |
P. tricornutum powder (0.5% or 1.0% (w/v)), gelatin solution |
Wound dressing with antimicrobial P. tricornutum-loaded gelatin nanofiber mat |
In vitro studies for antibacterial activity |
[72,73] |
5. Conclusions
Microalgae are constantly exposed to UV rays because they are usually found in areas with high sunlight exposure. Several studies have addressed the harmful impact of increased UV radiation on various microalgae. However, some microalgae have a great ability to overcome or counteract different stresses such as intense solar UV radiation, this has been happening over the years and with the evolution of microalgae, some examples of defense and/or tolerance mechanisms are: DNA repair, synthesis of antioxidants, and enzymatic/non-enzymatic compounds, such as MAAs and scytonemin to counteract the harmful effects of UV radiation. This characteristic has not only been investigated to learn more about microalgae, but also for applications such as product creation, since they grow rapidly and are considered the most promising and sustainable sources of biomass. Microalgae have shown a promising future in different areas of industry such as agriculture, energy, and cosmetics. Due to their performance under optimal conditions, they have been widely studied for different bioproducts. Bio-carriers can provide added value for the development of drug delivery systems in different microalgae bioproducts due to their benefits such as bioavailability, biodegradability, stability, availability, renewability, and low toxicity. To prevent photodamage, organisms have evolved to provide biochemical and physical defenses to UV by synthesizing robust bioactive compounds. For instance, phenylpropanoids and flavonoids in higher plants, melanin in animals, fungi, and microorganisms, and mycosporine-like amino acids (MAAs) in cyanobacteria, fungi, algae, and animals [
18,
19,
20]. Some algae species synthesize substances with chemical structures that can absorb UV radiation as well as inhibit actions on melanin synthesis [
19]. Studies have been performed regarding how UV radiation affects marine organisms; these studies concluded that different species of microalgae show a wide range of susceptibility to UV radiation [
20]. Some marine organisms, including microalgae, have developed physiological and biochemical traits to tolerate and cope with UV rays. Microalgae are considered the most important biomass producers on a global scale as they have rapid growth, easy to cultivate, and can tolerate a broad range of pH, salinity, and temperature [
23]. The UV radiation resistance capacity of microalgae and their potential applications in the field of sunscreens have been investigated, as they show an excellent capacity for topical application, meeting the growing demand for the creation of more environmentally friendly products.