Allergic diseases now affect over 30% of individuals in many communities, particularly young children, underscoring the need for effective prevention strategies in early life. These allergic conditions have been linked to environmental and lifestyle changes driving the dysfunction of three interdependent biological systems: microbiota, epithelial barrier and immune system. While this is multifactorial, dietary changes are of particular interest in the altered establishment and maturation of the microbiome, including the associated profile of metabolites that modulate immune development and barrier function. Prebiotics are non-digestible food ingredients that beneficially influence the health of the host by 1) acting as a fermentable substrate for some specific commensal host bacteria leading to the release of short-chain fatty acids in the gut intestinal tract influencing many molecular and cellular processes; 2) acting directly on several compartments and specifically on different patterns of cells (epithelial and immune cells). Nutrients with prebiotic properties are therefore of central interest in allergy prevention for their potential to promote a more tolerogenic environment through these multiple pathways.
- prebiotics
- mechanisms
- allergy
- microbiota
- immune system
- epithelial barrier
1. Introduction
2. Prebiotics
2.1. First Generation of Prebiotics
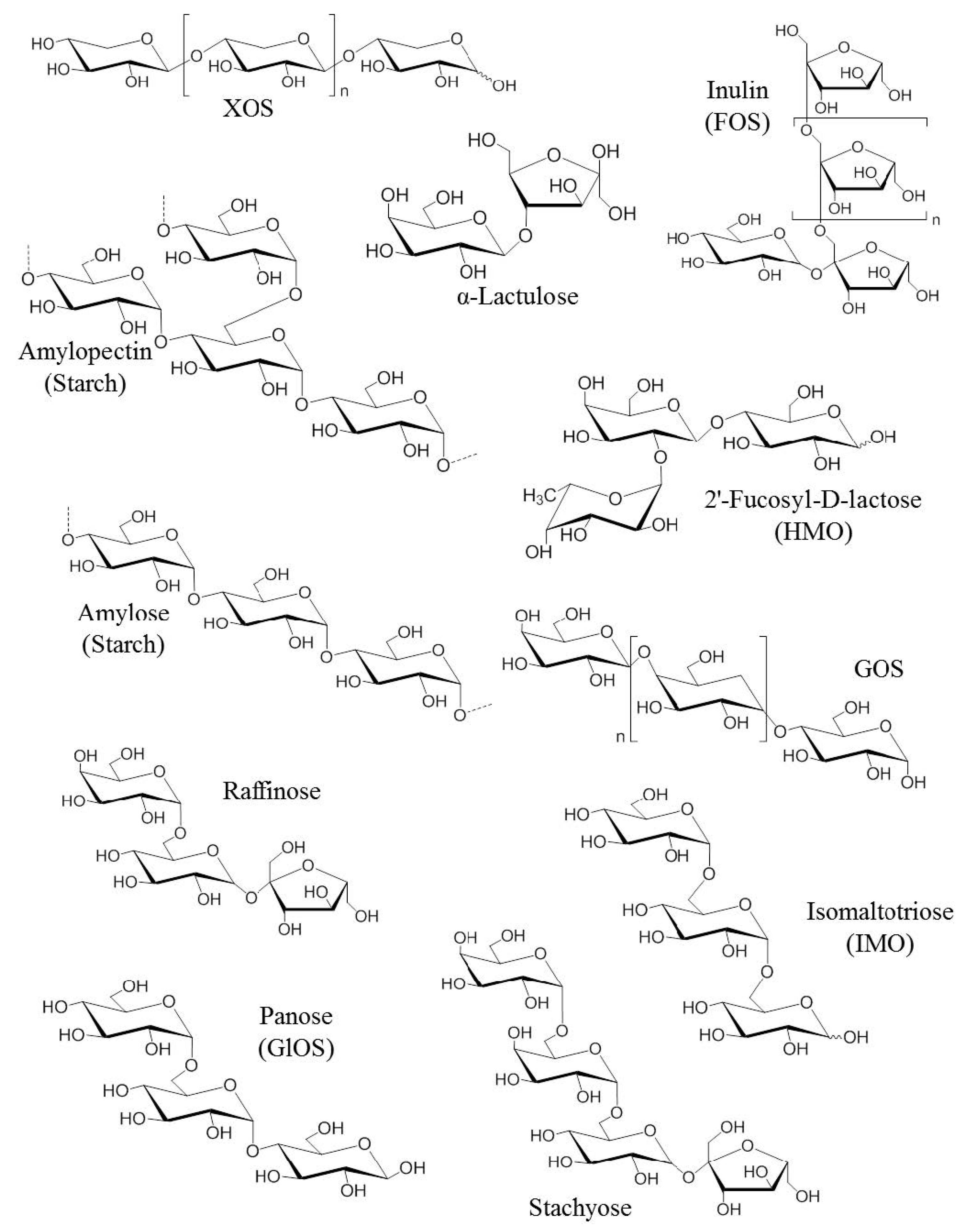
2.2. Second Generation of Prebiotics
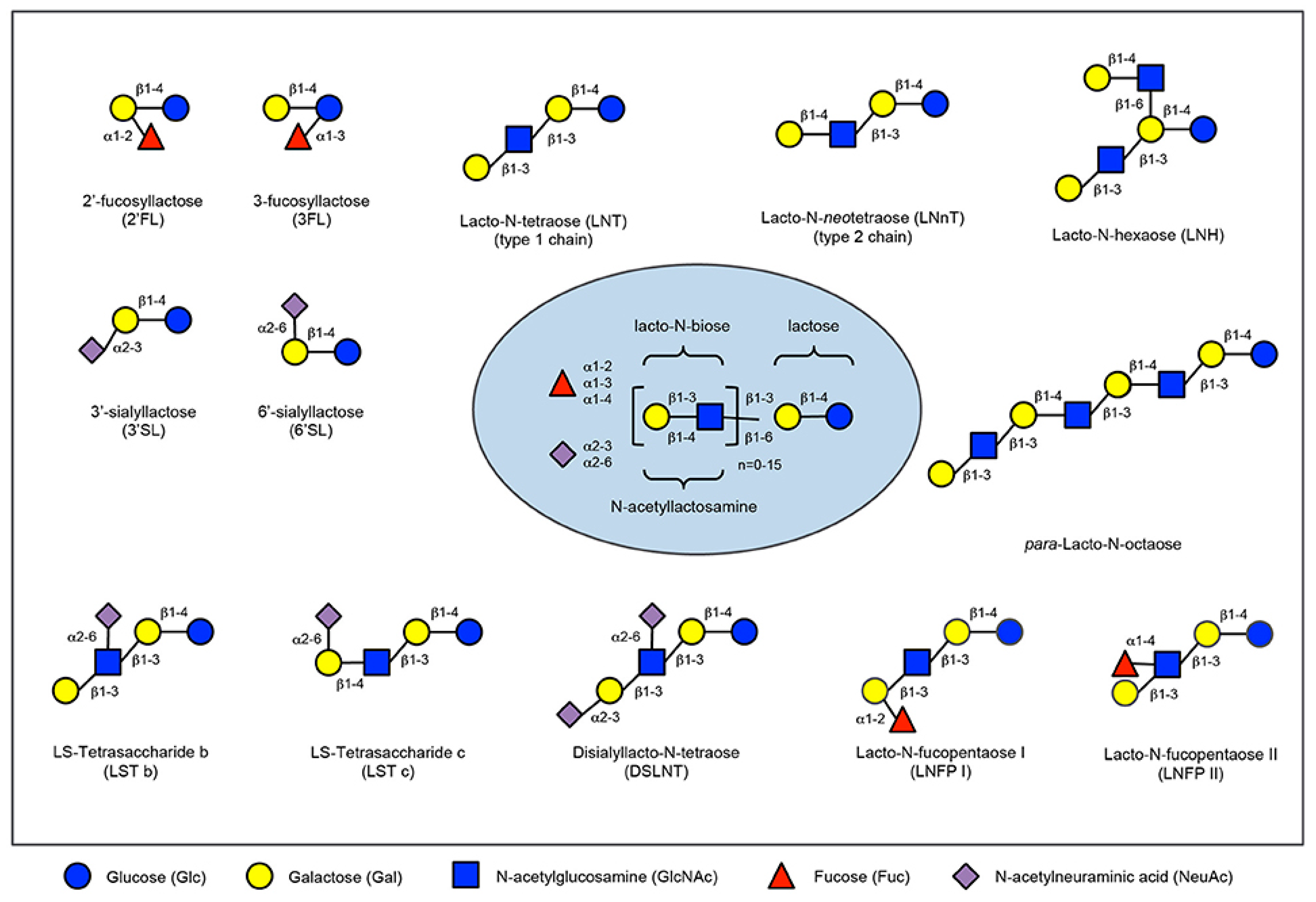
2.3. Human Milk Oligossacharides
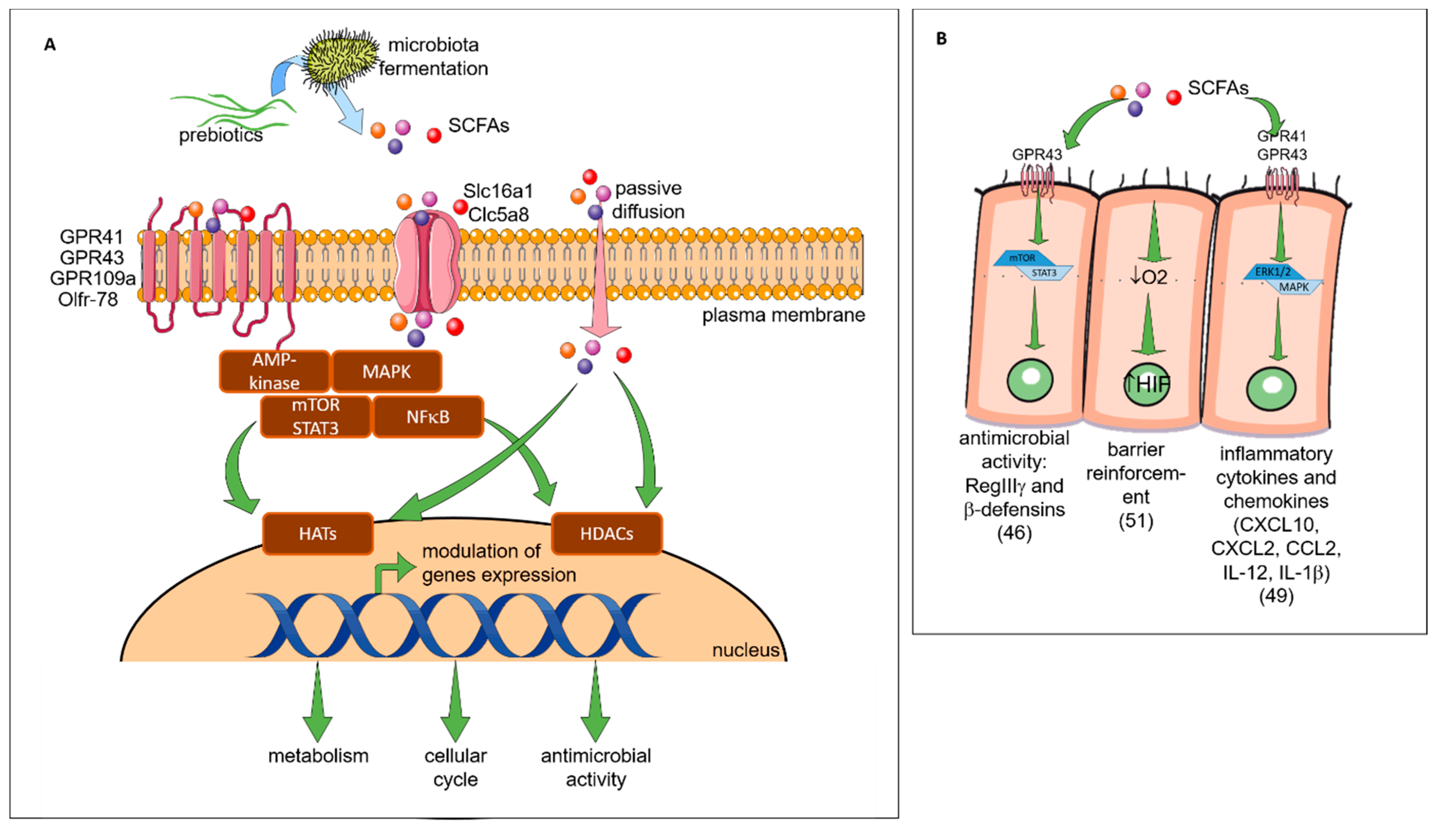
2.4. Prebiotics Mechanisms
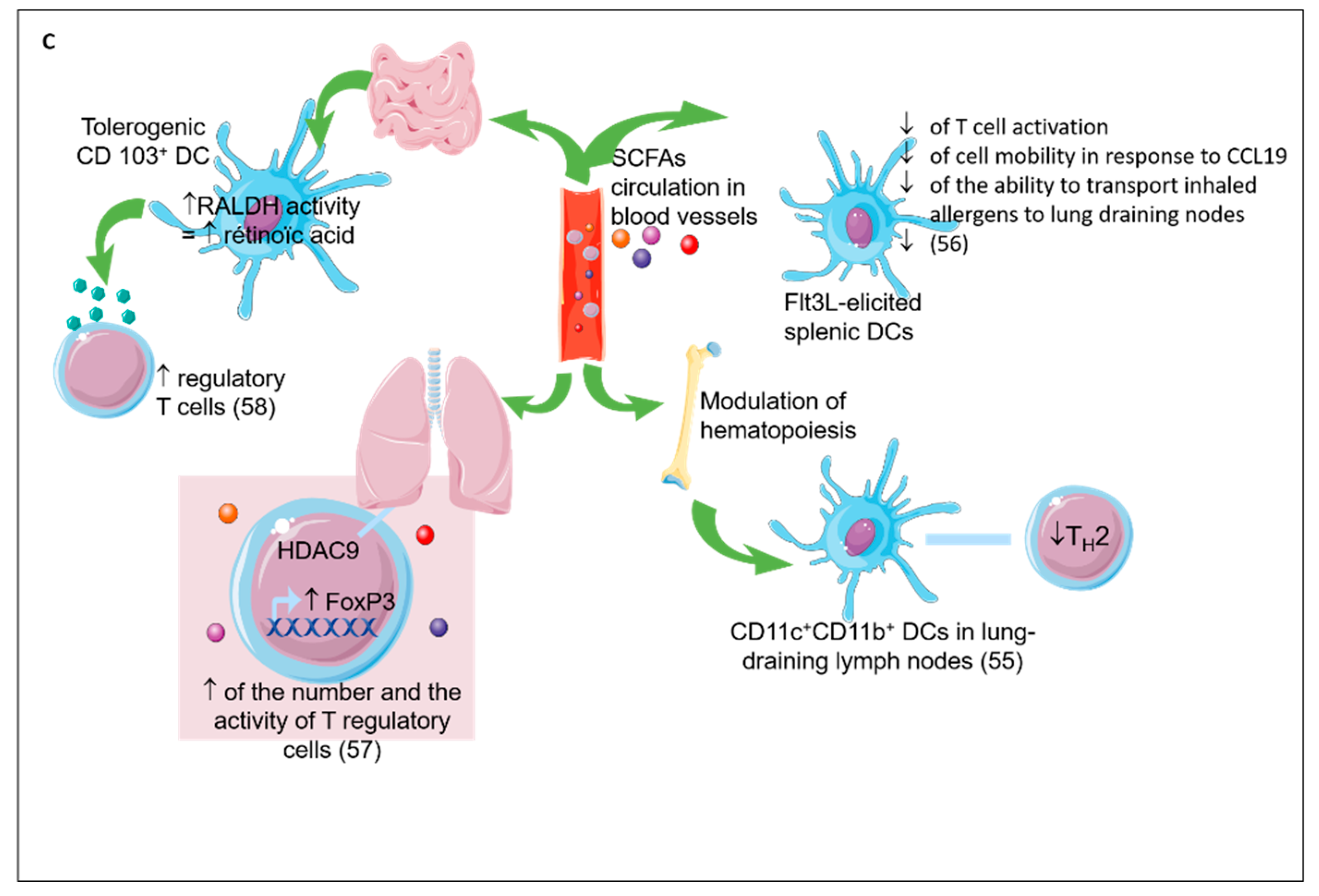
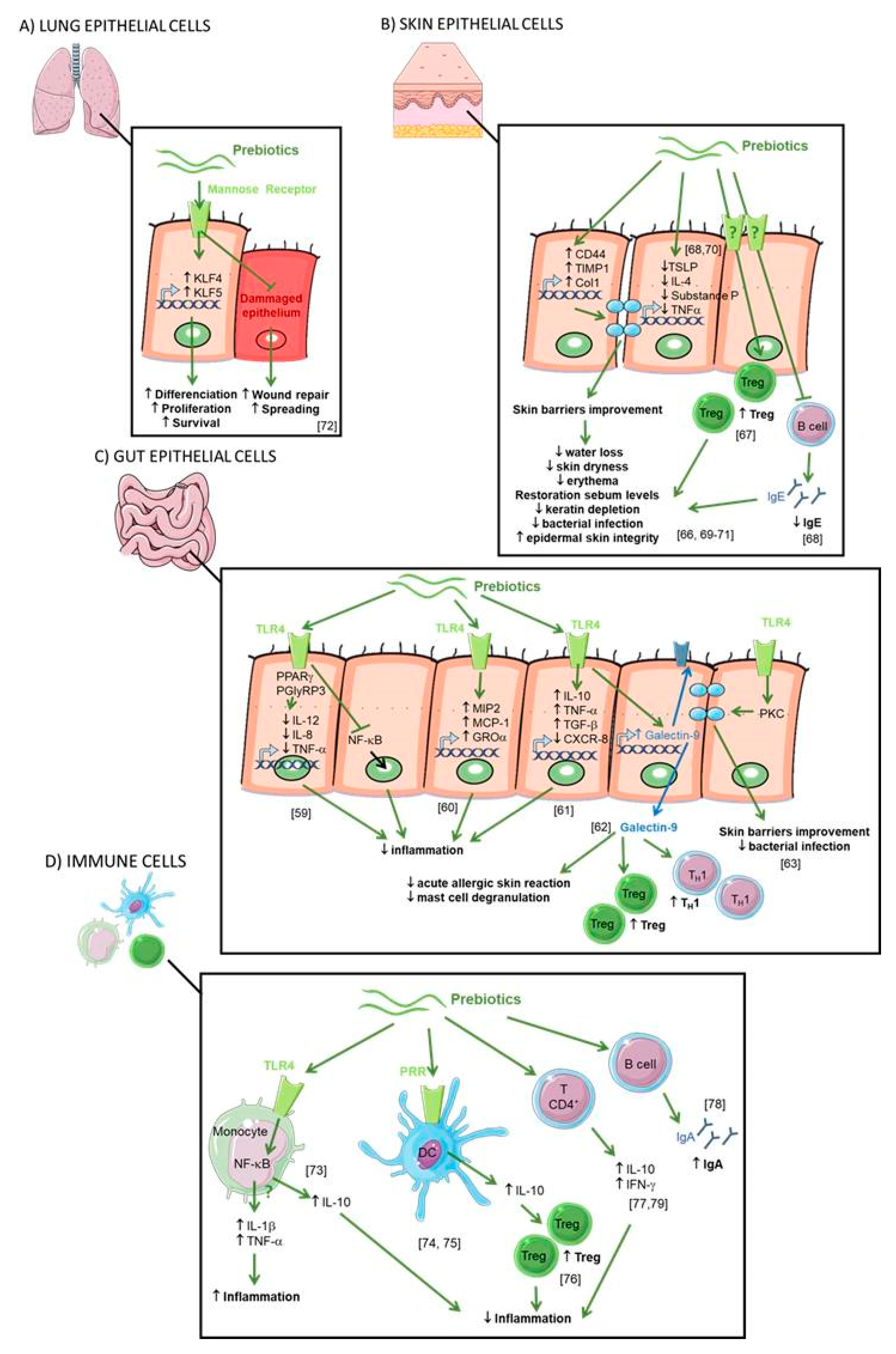
2.5. Direct Effect of Prebiotics (see Figure 4)
2.6. IEC
2.7. Skin Epithelial Cells
2.8. Lung Epithelial Cells
2.9. Immune System
This entry is adapted from the peer-reviewed paper 10.3390/nu11081841
References
- Prescott, S.L. Early-life environmental determinants of allergic diseases and the wider pandemic of inflammatory noncommunicable diseases. J. Allergy Clin. Immunol. 2013, 131, 23–30.
- Pawankar, R.; Canonica, G.W.; Holgate, S.T.; Lockey, R. World Allergy Organization (WAO) white book on allergy; World Allergy Organization: Milwaukee, WI, USA, 2011.
- Chinthrajah, R.S.; Hernandez, J.D.; Boyd, S.D.; Galli, S.J.; Nadeau, K.C. Molecular and cellular mechanisms of food allergy and food tolerance. J. Allergy Clin. Immunol. 2016, 137, 984–997.
- Nylund, L.; Nermes, M.; Isolauri, E.; Salminen, S.; De Vos, W.M.; Satokari, R. Severity of atopic disease inversely correlates with intestinal microbiota diversity and butyrate-producing bacteria. Allergy Eur. J. Allergy Clin. Immunol. 2015, 70, 241–244.
- Werfel, T.; Allam, J.P.; Biedermann, T.; Eyerich, K.; Gilles, S.; Guttman-Yassky, E.; Hoetzenecker, W.; Knol, E.; Simon, H.U.; Wollenberg, A.; et al. Cellular and molecular immunologic mechanisms in patients with atopic dermatitis. J. Allergy Clin. Immunol. 2016, 138, 336–349.
- West, C.E.; Jenmalm, M.C.; Prescott, S.L. The gut microbiota and its role in the development of allergic disease: A wider perspective. Clin. Exp. Allergy 2015, 45, 43–53.
- Chiu, C.Y.; Chan, Y.L.; Tsai, M.H.; Wang, C.J.; Chiang, M.H.; Chiu, C.C. Gut microbial dysbiosis is associated with allergen-specific IgE responses in young children with airway allergies. World Allergy Organ. J. 2019, 12, 1–12.
- Fieten, K.B.; Totté, J.E.E.; Levin, E.; Reyman, M.; Meijer, Y.; Knulst, A.; Schuren, F.; Pasmans, S.G.M.A. Fecal microbiome and food allergy in pediatric atopic dermatitis: A cross-sectional pilot study. Int. Arch. Allergy Immunol. 2018, 175, 77–84.
- Zimmermann, P.; Messina, N.; Mohn, W.W.; Finlay, B.B.; Curtis, N. Association between the intestinal microbiota and allergic sensitization, eczema, and asthma: A systematic review. J. Allergy Clin. Immunol. 2019, 143, 467–485.
- Demirci, M.; Tokman, H.; Uysa, H.; Demirvas, S.; Karakullukcu, A.; Saribas, S.; Cokugras, H.; Kocazeybey, B. Reduced Akkermansia muciniphila and Faecalibacterium prausnitzii levels in the gut microbiota of children with allergic asthma. Allergol. Immunopathol. 2019, 19, 1–10.
- Chassaing, B.; Vijay-kumar, M.; Gewirtz, A.T. How diet can impact gut microbiota to promote or endanger health. Curr. Opin. Gastroenterol. 2017, 33, 417–421.
- Tulic, M.K.; Hodder, M.; Forsberg, A.; McCarthy, S.; Richman, T.; D’Vaz, N.; Van Den Biggelaar, A.; Thornton, C.A.; Prescott, S.L. Differences in innate immune function between allergic and nonallergic children: New insights into immune ontogeny. J. Allergy Clin. Immunol. 2011, 127, 470–478.
- Tulic, M.K.; Andrews, D.; Crook, M.L.; Charles, A.; Tourigny, M.R.; Moqbel, R.; Prescott, S.L. Changes in thymic regulatory T-cell maturation from birth to puberty: Differences in atopic children. J. Allergy Clin. Immunol. 2012, 129, 199–206.
- Savage, J.; Johns, C.B. Food Allergy: Epidemiology and Natural History. Immunol Allergy Clin North. Am. 2015, 35, 45–59.
- Gollwitzer, E.S.; Marsland, B.J. Impact of Early-Life Exposures on Immune Maturation and Susceptibility to Disease. Trends Immunol. 2015, 36, 684–696.
- Renz, H.; Holt, P.G.; Inouye, M.; Logan, A.C.; Prescott, S.L.; Sly, P.D. An exposome perspective: Early-life events and immune development in a changing world. J. Allergy Clin. Immunol. 2017, 140, 24–40.
- Gibson, G.R.; Roberfroid, M.B. Dietary Modulation of the Human Colonie Microbiota: Introducing the Concept of Prebiotics. J. Nutr. 1995, 125, 1401–1412.
- Gibson, G.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.; Cani, P.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502.
- Kaczmarczyk, M.M.; Miller, M.J.; Freund, G.G. The health benefits of dietary fiber: beyond the usual suspects of type 2 diabetes, cardiovascular disease and colon cancer. Metabolism 2013, 61, 1058–1066.
- Bindels, L.B.; Delzenne, N.M.; Cani, P.D.; Walter, J. Opinion: Towards a more comprehensive concept for prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 303–310.
- Loo, J.V.; Coussement, P.; De Leenheer, L.; Hoebreg, H.; Smits, G. On the Presence of Inulin and Oligofructose as Natural Ingredients in the Western Diet. Crit. Rev. Food Sci. Nutr. 1995, 35, 525–552.
- Fernández, J.; Redondo-Blanco, S.; Miguélez, E.M.; Villar, C.J.; Clemente, A.; Lombó, F. Healthy effects of prebiotics and their metabolites against intestinal diseases and colorectal cancer. AIMS Microbiol. 2015, 1, 48–71.
- Depeint, F.; Tzortzis, G.; Vulevic, J.; I’anson, K.; Gibson, G.R. Prebiotic evaluation of a novel galactooligosaccharide mixture produced by the enzymatic activity of Bifidobacterium bifidum NCIMB 41171, in healthy humans: a randomized, double-blind, crossover, placebo-controlled intervention study. Am. J. Clin. Nutr. 2008, 87, 785–791.
- Vulevic, J.; Drakoularakou, A.; Yaqoob, P.; Tzortzis, G.; Gibson, G.R. Modulation of the fecal microflora profile and immune function by a novel trans-galactooligosaccharide mixture (B-GOS) in healthy elderly volunteers. Am. J. Clin. Nutr. 2008, 88, 1438–1446.
- Vulevic, J.; Juric, A.; Tzortzis, G.; Gibson, G.R. A Mixture of trans-Galactooligosaccharides Reduces Markers of Metabolic Syndrome and Modulates the Fecal Microbiota and Immune Function of Overweight Adults. J. Nutr. 2013, 143, 324–331.
- Silk, D.B.A.; Davis, A.; Vulevic, J.; Tzortzis, G.; Gibson, G.R. Clinical trial: The effects of a trans-galactooligosaccharide prebiotic on faecal microbiota and symptoms in irritable bowel syndrome. Aliment. Pharmacol. Ther. 2009, 29, 508–518.
- Searle, L.E.J.; Best, A.; Nunez, A.; Salguero, F.J.; Johnson, L.; Weyer, U.; Dugdale, A.H.; Cooley, W.A.; Carter, B.; Jones, G.; et al. A mixture containing galactooligosaccharide, produced by the enzymic activity of bifidobacterium bifidum, reduces salmonella enterica serovar typhimurium infection in mice. J. Med. Microbiol. 2009, 58, 37–48.
- Searle, L.E.J.; Cooley, W.A.; Jones, G.; Nunez, A.; Crudgington, B.; Weyer, U.; Dugdale, A.H.; Tzortzis, G.; Collins, J.W.; Woodward, M.J.; et al. Purified galactooligosaccharide, derived from a mixture produced by the enzymic activity of Bifidobacterium bifidum, reduces Salmonella enterica serovar Typhimurium adhesion and invasion in vitro and in vivo. J. Med. Microbiol. 2010, 59, 1428–1439.
- Drakoularakou, A.; Tzortzis, G.; Rastall, R.A.; Gibson, G.R. A double-blind, placebo-controlled, randomized human study assessing the capacity of a novel galacto-oligosaccharide mixture in reducing travellers’ diarrhoea. Eur. J. Clin. Nutr. 2010, 64, 146–152.
- Zivkovic, A.M.; German, J.B.; Lebrilla, C.B.; Mills, D.A. Human milk glycobiome and its impact on the infant gastrointestinal microbiota. Proc. Natl. Acad. Sci. USA 2010, 108, 4653–4658.
- Doherty, A.M.; Lodge, C.J.; Dharmage, S.C.; Dai, X.; Bode, L.; Lowe, A.J. Human Milk Oligosaccharides and Associations with immune-mediated disease and infection in childhood: A systematic review. Front. Pediatr. 2018, 6, 1–8.
- Kunz, C.; Rudloff, S. Compositional analysis and metabolism of human milk oligosaccharides in infants. Nestle Nutr. Inst. Workshop Ser. 2017, 88, 137–147.
- Donovan, S.M.; Comstock, S.S. Human milk oligosaccharides influence neonatal mucosal and systemic immunity. Ann. Nutr. Metab. 2016, 69, 42–51.
- Wang, M.; Li, M.; Wu, S.; Lebrilla, C.B.; Chapkin, R.S.; Ivanov, I.; Donovan, S.M. Fecal microbiota composition of breast-fed infants is correlated with human milk oligosaccharides consumed. J. Pediatr. Gastroenterol. Nutr. 2015, 60, 825–833.
- Triantis, V.; Bode, L.; van Neerven, R.J.J. Immunological Effects of Human Milk Oligosaccharides. Front. Pediatr. 2018, 6, 1–14.
- Slavin, J. Fiber and prebiotics: Mechanisms and health benefits. Nutrients 2013, 5, 1417–1435.
- Vandeputte, D.; Falony, G.; Vieira-Silva, S.; Wang, J.; Sailer, M.; Theis, S.; Verbeke, K.; Raes, J. Prebiotic inulin-type fructans induce specific changes in the human gut microbiota. Gut 2017, 66, 1968–1974.
- Hopkins, M.J.; Macfarlane, G.T. Nondigestible oligosaccharides enhance bacterial colonization resistance against Clostridium difficile in vitro. Appl. Environ. Microbiol. 2003, 69, 1920–1927.
- Fukuda, S.; Toh, H.; Hase, K.; Oshima, K.; Nakanishi, Y.; Yoshimura, K.; Tobe, T.; Clarke, J.M.; Topping, D.L.; Suzuki, T.; et al. Bifidobacteria can protect from enteropathogenic infection through production of acetate. Nature 2011, 469, 543–549.
- Tochio, T.; Kadota, Y.; Tanaka, T.; Koga, Y. 1-Kestose, the smallest fructooligosaccharide component, which efficiently stimulates faecalibacterium prausnitzii as well as bifidobacteria in humans. Foods 2018, 7, 140.
- Chung, W.S.F.; Meijerink, M.; Zeuner, B.; Holck, J.; Louis, P.; Meyer, A.S.; Wells, J.M.; Flint, H.J.; Duncan, S.H. Prebiotic potential of pectin and pectic oligosaccharides to promote anti-inflammatory commensal bacteria in the human colon. FEMS Microbiol. Ecol. 2017, 93, 1–9.
- Corrêa-Oliveira, R.; Fachi, J.L.; Vieira, A.; Sato, F.T.; Vinolo, M.A.R. Regulation of immune cell function by short-chain fatty acids. Clin. Transl. Immunol. 2016, 5, 1–8.
- Hiroki, T.; Tohru, Y.; Ryo, I.; Kazunari, U.; Kumiko, T.; Junko, N. The cellular expression of SMCT2 and its comparison with other transporters for monocarboxylates in the mouse digestive tract. Biomed. Res. 2010, 31, 239–249.
- Takebe, J.; Nio, J.N.; Orimatsu, M.M.; Araki, S.K.; Uwahara, A.K.; Ato, I.K. Histochemical demonstration of a Na+ -coupled transporter for short-chain fatty acids (Slc5a8) in the intestine and kidney of the mouse. Biomed. Res. 2005, 26, 213–221.
- Den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.-J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340.
- Jung, T.H.; Park, J.H.; Jeon, W.M.; Han, K.S. Butyrate modulates bacterial adherence on LS174T human colorectal cells by stimulating mucin secretion and MAPK signaling pathway. Nutr. Res. Pract. 2015, 9, 343–349.
- Zhao, Y.; Chen, F.; Wu, W.; Sun, M.; Bilotta, A.J.; Yao, S.; Xiao, Y.; Huang, X.; Eaves-Pyles, T.D.; Golovko, G.; et al. GPR43 mediates microbiota metabolite SCFA regulation of antimicrobial peptide expression in intestinal epithelial cells via activation of mTOR and STAT3. Mucosal Immunol. 2018, 11, 752–762.
- Li, M.; Van Esch, B.C.A.M.; Henricks, P.A.J.; Garssen, J.; Folkerts, G. Time and Concentration Dependent Effects of Short Chain Fatty Acids on Lipopolysaccharide- or Tumor Necrosis Factor α -Induced Endothelial Activation. Front. Pharmacol. 2018, 9, 1–12.
- Li, M.; Van Esch, B.C.A.M.; Wagenaar, G.T.M.; Garssen, J.; Folkerts, G.; Henricks, P.A.J. Pro- and anti-inflammatory effects of short chain fatty acids on immune and endothelial cells. Eur. J. Pharmacol. 2018, 831, 52–59.
- Zenhom, M.; Hyder, A.; de Vrese, M.; Heller, K.J.; Roeder, T.; Schrezenmeir, J. Prebiotic Oligosaccharides Reduce Proinflammatory Cytokines in Intestinal Caco-2 Cells via Activation of PPAR and Peptidoglycan Recognition Protein 3. J. Nutr. 2011, 141, 971–977.
- Ortega-González, M.; Ocón, B.; Romero-Calvo, I.; Anzola, A.; Guadix, E.; Zarzuelo, A.; Suárez, M.D.; Sánchez de Medina, F.; Martínez-Augustin, O. Nondigestible oligosaccharides exert nonprebiotic effects on intestinal epithelial cells enhancing the immune response via activation of TLR4-NFκB. Mol. Nutr. Food Res. 2014, 58, 384–393.
- Johnson-Henry, K.C.; Pinnell, L.J.; Waskow, A.M.; Irrazabal, T.; Martin, A.; Hausner, M.; Sherman, P.M. Short-Chain Fructo-oligosaccharide and Inulin Modulate Inflammatory Responses and Microbial Communities in Caco2-bbe Cells and in a Mouse Model of Intestinal Injury. J. Nutr. 2014, 144, 1725–1733.
- De Kivit, S.; Saeland, E.; Kraneveld, A.D.; Van De Kant, H.J.G.; Schouten, B.; Van Esch, B.C.A.M.; Knol, J.; Sprikkelman, A.B.; Van Der Aa, L.B.; Knippels, L.M.J.; et al. Galectin-9 induced by dietary synbiotics is involved in suppression of allergic symptoms in mice and humans. Allergy 2012, 67, 343–352.
- Wu, R.Y.; Abdullah, M.; Määttänen, P.; Pilar, A.V.C.; Scruten, E.; Johnson-Henry, K.C.; Napper, S.; O’Brien, C.; Jones, N.L.; Sherman, P.M. Protein kinase C δ signaling is required for dietary prebiotic-induced strengthening of intestinal epithelial barrier function. Sci. Rep. 2017, 7, 1–10.
- Réquilé, M.; Gonzàlez Alvarez, D.O.; Delanaud, S.; Rhazi, L.; Bach, V.; Depeint, F.; Khorsi-Cauet, H. Use of a combination of in vitro models to investigate the impact of chlorpyrifos and inulin on the intestinal microbiota and the permeability of the intestinal mucosa. Environ. Sci. Pollut. Res. 2018, 25, 1–12.
- Bermudez-Brito, M.; Sahasrabudhe, N.M.; Rösch, C.; Schols, H.A.; Faas, M.M.; De Vos, P. The impact of dietary fibers on dendritic cell responses in vitro is dependent on the differential effects of the fibers on intestinal epithelial cells. Mol. Nutr. Food Res. 2015, 59, 698–710.
- Hong, K.B.; Jeong, M.; Han, K.S.; Hwan Kim, J.; Park, Y.; Suh, H.J. Photoprotective effects of galacto-oligosaccharide and/or Bifidobacterium longum supplementation against skin damage induced by ultraviolet irradiation in hairless mice. Int. J. Food Sci. Nutr. 2015, 66, 923–930.
- Kim, H.J.; Lee, S.H.; Go, H.N.; Ahn, J.R.; Kim, H.J.; Hong, S.J. Effects of kestose on gut mucosal immunity in an atopic dermatitis mouse model. J. Dermatol. Sci. 2018, 89, 27–32.
- Suzuki, H.; Oomizu, S.; Yanase, Y.; Onishi, N.; Uchida, K.; Mihara, S.; Ono, K.; Kameyoshi, Y.; Hide, M. Hydrolyzed Konjac glucomannan suppresses IgE production in mice B cells. Int. Arch. Allergy Immunol. 2010, 152, 122–130.
- Onishi, N.; Kawamoto, S.; Ueda, K.; Yamanaka, Y.; Katayama, A.; Suzuki, H.; Aki, T.; Hashimoto, K.; Hide, M.; Ono, K. Dietary Pulverized Konjac Glucomannan Prevents the Development of Allergic Rhinitis-Like Symptoms and IgE Response in Mice. Biosci. Biotechnol. Biochem. 2007, 71, 2551–2556.
- Hong, Y.H.; Chang, U.J.; Kim, Y.S.; Jung, E.Y.; Suh, H.J. Dietary galacto-oligosaccharides improve skin health: A randomized double blind clinical trial. Asia Pac. J. Clin. Nutr. 2017, 26, 613–618.
- Narbutt, J.; Bednarski, I.A.; Lesiak, A. The effect of an emollient with benfothiamine and Biolin prebiotic on the improvement of epidermal skin function. Postep. Dermatologii i Alergol. 2016, 33, 224–231.
- Smith, A.R.; Knaysi, G.; Wilson, J.M.; Wisniewski, J.A. The Skin as a Route of Allergen Exposure: Part I. Immune Components. Curr Allergy Asthma Rep. 2017, 17, 1–18.
- Michael, C.F.; Waters, C.M.; LeMessurier, K.S.; Samarasinghe, A.E.; Song, C.Y.; Malik, K.U.; Lew, D.B. Airway Epithelial Repair by a Prebiotic Mannan Derived from Saccharomyces cerevisiae. J. Immunol. Res. 2017, 2017, 1–7.
- Capitan-Canadas, F.; Ortega-Gonzalez, M.; Guadix, E.; Zarzuelo, A.; Suarez, M.D.; de Medina, F.S.; Martinez-Augustin, O. Prebiotic oligosaccharides directly modulate proinflammatory cytokine production in monocytes via activation of TLR4. Mol. Nutr. Food Res. 2014, 58, 1098–1110.
- De Kivit, S.; Kraneveld, A.D.; Garssen, J.; Willemsen, L.E.M. Glycan recognition at the interface of the intestinal immune system: Target for immune modulation via dietary components. Eur. J. Pharmacol. 2011, 668, 124–132.
- Lehmann, S.; Hiller, J.; Van Bergenhenegouwen, J.; Knippels, L.M.J.; Garssen, J.; Traidl-Hoffmann, C. In vitro evidence for immune-modulatory properties of non-digestible oligosaccharides: Direct effect on human monocyte derived dendritic cells. PLoS ONE 2015, 10, 1–15.
- Perdijk, O.; Joost van Neerven, R.J.; Van den Brink, E.; Savelkoul, H.F.J.; Brugman, S. The oligosaccharides 6′-sialyllactose, 2′-fucosyllactose or galactooligosaccharides do not directly modulate human dendritic cell differentiation or maturation. PLoS One 2018, 13, 1–15.
- Roller, M.; Rechkemmer, G.; Watzl, B. Prebiotic Inulin Enriched with Oligofructose in Combination with the Probiotics Lactobacillus rhamnosus and Bifidobacterium lactis Modulates Intestinal Immune Functions in Rats. J. Nutr. 2018, 134, 153–156.
- Ito, H.; Takemura, N.; Sonoyama, K.; Kawagishi, H.; Topping, D.L.; Conlon, M.A.; Morita, T. Degree of polymerization of inulin-type fructans differentially affects number of lactic acid bacteria, intestinal immune functions, and immunoglobulin a secretion in the rat cecum. J. Agric. Food Chem. 2011, 59, 5771–5778.
- Takahashi, T.; Nakagawa, E.; Nara, T.; Yajima, T.; Kuwata, T. Effects of Orally Ingested Bifidobacterium longum on the Mucosal IgA Response of Mice to Dietary Antigens. Biosci. Biotechnol. Biochem. 2005, 62, 10–15.