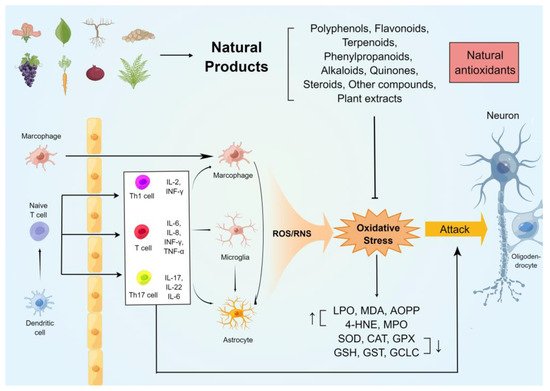
2. MS 动物模型中抗氧化应激的天然化合物
近年来,天然产物在 MS 中的抗氧化作用得到了广泛的研究。天然产物已用于多发性硬化的啮齿动物模型,并被发现可减少中枢神经系统和外周免疫炎症反应,减轻脱髓鞘和轴突变性,并减轻大脑、脊髓和视神经的氧化应激损伤,从而改善各种临床症状女士。这些天然产物在 MS 动物模型(大鼠和小鼠)中的抗氧化和神经保护作用机制总结在 表 1中。
表 1. MS 动物模型中使用的天然产物的抗氧化应激特性。
| 化合物 |
实验模型 |
治疗时间点及路径 |
剂量 |
抗氧化应激作用 |
主要成果 |
| 花青素 |
EB
(雄性 Wistar 大鼠) |
从 EB 注射日
(口服)开始 |
30, 100 mg/kg 持续 7 天 |
↑ HNE-His、MDA、NOX
↑ NPSH、GSH、SOD |
↓ IL-6、IL-1β、TNF-α、IFN-γ。
↑ IL-10。
↑ Na +、K + -ATP酶、Ca2+-ATP酶。 |
| 橙皮苷 |
EAE
(雌性 C57BL/6J 小鼠) |
在第 14 天诱导 EAE
(腹腔注射)后 |
100 毫克/公斤/天,持续 7 天 |
↓ MDA。
↑ GPx、SOD、CAT、GSH |
↓ Caspase-3、IL-17、TNF-α 和 IL-1β。
↓ 炎症浸润。 |
| 甘草查耳酮 A |
EAE
(雌性 C57BL/6 小鼠) |
从免疫后第 10 天开始
(口服) |
15、30 毫克/公斤/天,持续 10 天 |
↓ H2O2, NO |
↓ TNF-α、IFN-γ 和 IL-17。
↓ Th1 和 Th17 细胞。 |
| 黄酮氧化物 |
EAE
(雌性 C57BL/6 小鼠) |
预防方案:从免疫日开始 治疗方案:从免疫后第 14 天开始
(腹腔注射) |
每隔一天100 mg/kg/d
预防方案:
持续 28 天。
治疗方案:19天。 |
↓ iNOS、COX2、5-LOX。 |
↓ IL-6、IL-12、IL-23、IL-17、IFN-γ。
↓ Th1/Th17 细胞。
↑ Arg1、Ym1、CD206、TGF-β、IL-10。 |
| 木犀草素 |
EAE
(雌性 Wistar 大鼠) |
从免疫后第 10 天开始
(腹腔注射) |
10 毫克/公斤/天,持续 33 天 |
↑ TAC |
↑ CNTF,营地。
↓ c-caspase-3、NF-κB、MIP-1α。 |
| 淫羊藿黄酮 |
EAE
(雌性 Lewis 大鼠) |
从免疫日开始
(口服) |
20、60 毫克/公斤/天
,持续 15 天 |
↓ 不,iNOS |
↓ IL-1β、TNF-α、IκB-α、Iba1、CD3、GFAP。
↑ CNP酶,NGF。 |
| 科拉维龙 |
CPZ
(male Wistar rats) |
Starting from cuprizone diet fed
(oral) |
200 mg/kg/d
both cuprizone and kolaviron for 42 days |
↓ MDA.
↑ GPx, SOD |
↑ the neuronal integrity |
| Quercetin |
EB
(male Wistar rats) |
Starting from EB injection day
(i.p. injection) |
50 mg/kg/d
for 22 days |
↓ MDA.
↑ CAT, SOD. |
↓ AChE.
↓ Na+, K+-ATPase |
| Phloretin |
EAE
(C57BL/6J mice) |
Starting after 6 days of immunization or after disease onset
(i.p. injection) |
50 mg/kg/d
for 10 days
or 20 days |
↓ ROS, NO, NOS2,
↑ Nrf2, NQO1 |
↓ MHC-II, CD86, TNF-α, IL-6, CCL4, CCL5, CXCL2.
↑ IL-4, CNTF, IGF-1, p-AMK, P62, LC3-II. |
| Curcumin |
EAE
(female C57BL/6 mice) |
Starting from immunization day
(i.p. injection) |
20 mg/kg/d
for 21 days |
↑ GPx, SOD. |
↓ IL-6, IL-17, TNF-α, IFN-γ.
↑ TGF-β. |
| |
EAE
(female Lewis rats) |
Starting at 12th day post-immunization |
12.5 mg/kg/d
for 18 days |
↓ iNOS, ↑ HO-1, Nrf2 |
↓ IL-1, IL-17.
↑ IL-4, IL-10, TGF-β, BDNF, NGF, MBP, Nestin. |
| Resveratrol |
EAE
(female C57BL/6 mice) |
Starting from immunization day
(i.p. injection) |
10, 25, 50 mg/kg/d
for 20 days |
↓ iNOS, NOX2, NOX4.
↓ NADPH activity |
↑ Arg1 and IL-10
↑ ZO-1, Occludin, claudin-5, ICAM-1, VCAM-1.
↑ the BBB integrity |
| |
CPZ
(male C57BL/6 mice) |
cuprizone diet for 7 days, followed by 3 weeks on 0.2 % cuprizone diet plus resveratrol
(oral) |
250 mg/kg/d
for 21 days |
↓ LPO. ↑ GSH, SOD, cytochrome oxidase |
↓ Rel-A, pIκB-α, TNF-α.
↑ MBP, CNP, Olig1. |
| Acteoside |
EAE
(female C57BL/6N mice) |
Prevention protocol: Starting from day 2 post immunization
therapeutic treatment protocol: from day 11 post immunization
(oral) |
Prevention protocol: 30 mg/kg/d for 29 days
therapeutic treatment protocol: 5, 10 and 30 mg/kg/d for 19 days |
↓ ONOO-, iNOS, NADPH oxidases. |
↓ LC3-II to LC3-I in mitochondrial fraction.
↓ the translocation of Drp1 to the mitochondria.
↓ neuronal apoptotic. |
| Oleacein |
EAE
(female C57BL/6J mice) |
Starting from immunization day
(i.p. injection) |
10 mg/kg/d
for 24 days |
↓ O2−, MDA, AOPP, ROS, iNOS, COX2.
↑ FRAP, Sestrin-3 |
↓ TNFα, IL-13, IL-33, IL-1β, MCP-1.
↑ IL-10.
↑ the BBB integrity.
↓ inflammatory infiltration. |
| Ellagic acid |
CPZ
(male C57BL/6 mice) |
Starting from cuprizone diet fed
(oral) |
5, 50, 100 mg/kg/d
coadministration of CPZ and Ellagic acid for 6 weeks |
↓ MDA, ROS.
↑ CAT, SOD. |
↑ the integrity of myelin in spinal cord and sciatica. |
| Arbutin |
LPC
(male Wistar rats) |
Starting from LPC injection day
(i.p. injection) |
50 mg/kg/d
for 14 days |
↑ Nrf2, HO-1. |
↓ IL-1β, IL-17, TNF-α, GFAP.
↑ IL-10, MBP, Olig2. |
| Paeonol |
CPZ
(male C57BL/6 mice) |
Started 1 week after beginning cuprizone challenge till the end of week 6 post-cuprizone
(oral) |
25 mg, 100 mg/kg/d
for 42 days |
↓ MDA, ROS, MPO.
↑ CAT, SOD, GSH. |
↓ NF-κB, TNF-α.
↑ MBP. |
| 18β-glycyrrhetinic acid |
EAE
(female C57BL/6 mice) |
14 days after induction of EAE
(i.p. injection) |
100 mg/kg/d
for 7 days |
↓ MDA. ↑ GPx, SOD, CAT, GSH |
↓ Caspase-3, IL-17, TNF-α, IL-1β. |
| β-Caryophyllene |
EAE
(female C57BL/6 mice) |
Starting at 10th day post-immunization
(oral) |
25, 50mg/kg/d
for 9 days |
↓ H2O2, NO |
↓ TNF-α, IFN-γ, IL-17 |
| Ginkgolide K |
CPZ
(male C57BL/6 mice) |
Starting from 5th weeks until the end of 6th weeks
(oral) |
20 mg/kg/d
for 14 days |
↑ Nrf2, HO-1.
↓ NO, iNOS. |
↓ IL-1β, IL-6, TNF-α.
↓ p-NF-kB/p65, caspase-3, TUNEL.
↑ IGF/PI3K. |
| Bixin |
EAE
(female C57BL/6 mice) |
Starting at 12th days post-immunization
(i.p. injection) |
50, 100, 200 mg/kg/d for 18 days |
↓ ROS, Nrf2, TXNIP, NLRP3 |
↓ TNF-α, IL-6, IL-8, IL-17, and IFN-γ.
↑ IL-10.
↓ Th1 and Th17 cells. |
| Acetyl-11-keto-β-boswellic acid |
EAE
(female SJL/J mice) |
starting at day 11 after immunization
(oral) |
20 mg/kg/d
for 30 days |
↑ Nrf2, HO-1, TAC
↓ iNOS, lipid peroxides |
↓ p-NF-κB, NF-κB, IL-6 |
| Ginsenoside-Rg3 |
EAE
(female C57BL/6N mice) |
Starting from 7 days before immunization
(oral) |
37.5, 75, 150mg/kg/d
for 28 day |
↓ MitoSOX, 4-HNE, NOX2, NOX4, NADPH activity, COX2, iNOS |
↑ occludens-1, claudin-3, and claudin-5.
↓ TNF-α, IL-6, IL-1β. |
| Astragaloside |
EAE
(female C57BL/6 mice) |
Starting from immunization day
(i.p. injection) |
10, 25, 50 mg/kg/d
for 15 days |
↓ ROS, MDA, iNOS
↑ GPx, SOD |
↓ p53, p-tau
↑ Bcl-2/Bax ratio.
↑ the BBB integrity. |
| Nordihydroguaiaretic acid |
EAE
(female C57BL/6 mice) |
Starting from immunization day
(i.p. injection) |
10 mg/kg/d
for 30 days |
↓ MDA. ↑ HO-1 |
↓ IL-17
↓ p38MAPK, SGK1.
↓ inflammatory infiltration. |
| Caffeic acid phenethyl ester |
EAE
(female Wistar rats) |
Starting from the first day of immunization
(i.p. injection) |
25 μmol/kg/d
for 14 days |
↑ GPx, ADA, SOD.
↓ MDA, NO, XO. |
↓ inflammatory infiltration. |
| P-Coumaric Acid |
CPZ
(female and male C57BL/6 mice) |
Starting from 7th weeks until the end of 12th weeks
(oral) |
21.6 ppm/mouse/d
for 42 days |
↓ free radical (DPPH) |
↓ MMP-9 |
| Magnolol |
EAE
(female Swiss mice) |
Starting from immunization day
(oral) |
0.1, 1, 10 mg/kg/d
for 21 days |
↓ MDA, MPO, NO, iNOS. ↑ GSH, GST, SOD, Nrf2. |
↓ CD8+ T cell.
↓ c-caspase-3. |
| Daphnetin |
EAE
(female C57BL/6 mice) |
Starting from immunization day
(i.p. injection) |
8 mg/kg/d
for 21 days |
↓MDA, HO-1 |
↓ IFN-γ, IL-17, IL-1β, IL-6, TNF-α.
↑ IL-10.
inflammatory infiltration. |
| Piperine |
EAE
(female Lewis rats) |
Starting at 8th day post-immunization
(i.p. injection) |
5 mg/kg/d
for 22 days |
↓ iNOS, MDA.
↑ Nrf2, HO-1
FRAP. |
↓ TNF-, IL-1β, caspase-3.
↑ IL-10, BDNF, MBP. |
| |
LPC
(Male Wistar rats) |
Starting at 3 days post
LPC injection
(i.p. injection) |
5, 10, 20 mg/kg/d
for 10 days |
↑ Nrf2, HO1, FRAP.
↓ iNOS. |
↓ TNF-α, IL-1β, NF-κB, Foxp3.
↑ IL-10, BDNF, MBP. |
| Matrine |
EAE
(Female Wistar rats) |
starting from day 11 post immunization
(i.p. injection) |
250 mg/kg/d
for 7 days |
↓ MDA. ↑ GPx |
↓ caspase-3, α-B-crystallin, Cyt.
↑ Beclin1, LC3. |
| Shikonin |
EAE
(female C57BL/6 mice) |
After EAE was induced
(i.p. injection) |
20 mg/kg/d
for 21 days |
↑ GPx1 |
↓ TNF-α, IFN-γ and Bax.
↑ TGF-β and Bcl2. |
| Thymoquinone |
EAE
(female Lewis rats) |
at days 1–5 post immunization
or
at day 12–17 post immunization
(i.v. injection) |
1 mg/kg
for 5 days or 6 days |
↑ GSH |
↓ IL-7, IL-7R
↓ inflammatory infiltration. |
| Withametelin |
EAE
(Female Swiss mice) |
starting at day 9 through day 25
(i.p. injection) |
10, 100, and 1000 μg/kg/d
for 17 days |
↓ NO, MPO, iNOS
↑ GSH, SOD, Nrf2, HO-1 |
↓ TLR4, NF-κB, AP-1.
↑ IκB-α, Bcl-2. |
| Guggulsterone |
EB
(Wistar rats) |
Starting at 8th day
post EB injection
(oral) |
30, 60 mg/kg/d
for 28 days |
↓ AchE, MDA, NO.
↑ SOD, CAT, GSH. |
↓ IL-1β, TNF-α.
↓ caspase-3, Bax, STAT3.
↑ Bcl2, PPAR-ϒ, MBP. |
| Sulforaphane |
EAE
(female C57BL/6N mice) |
14 days before
EAE induction
(oral) |
50 mg/kg/d
for 28 days |
↓ iNOS |
↓ CD4, CD68, GFAP
↓ inflammatory infiltration. |
| |
EAE
(female C57BL/6 mice) |
Starting from immunization day
(i.p. injection) |
50 mg/kg
every other day up to 22 days |
↓ HO-1, NQO1
↑ Nrf2 |
↑ Occludin, claudin-5. ↑IL-10.
↓ MMP9, Th17 cells. |
| 3H-1,2-dithiole-3-thione |
EAE
(female C57BL/6N mice) |
starting at day 1 post immunization
(i.p. injection) |
10 mg/kg/d
for 30 days |
↑ Nrf2, NQO1, GCLC, HO-1.
↓ iNOS |
↓ IL-23.
↓ Th1 and Th17 differentiation. |
| C-Phycocyanin |
EAE
(female C57BL/6 mice) |
Treatments started at disease onset (i.p. injection) |
2, 4, 8 mg/kg/d
for 15 days |
↓ MDA, peroxidation potential, CAT/SOD ratio. ↑ GSH. |
↓ CD3, Mac-3
↓ IL-17, IL-6, Foxp3
↓ inflammatory infiltration |
| Artemisia dracunculus L. |
EAE
(female C57BL/6 mice) |
Starting at 11th day post immunization
(oral) |
500 mg/kg/d
for 23 days |
↑ TAC
(FRAP) |
↓ IL-17, IL-23.
↑ TGF-β.
↓ inflammatory infiltration |
| Olive leaf |
EAE
(female C57BL/6 mice) |
starting from the first day after EAE induction.
(olive leaf tea, oral)
starting from the 8th day after EAE induction
(olive leaf extract, i.p. injection) |
Olive leaf tea: ad libitum for 20 or 30 days
Olive leaf extract: 1024 mg/kg/d
for 10 days |
↓ MDA
↑ SOD1, SOD2, GPx1 |
↑ SIRT.
↓ M1 microglia.
↑ M2 microglia. |
| Melilotus officinalis |
EAE
(female C57BL/6 mice) |
starting from first day post-immunization
(i.p. injection) |
10 mg/kg/d
for 21 days |
↑ CAT, GPx1 |
↓ IL-6, TNF-α, IFN-γ, IL-17.
↑ TGF-β, IL-5. |
| Nutshell of Xanthoceras sorbifolia |
EAE
(female C57BL/6 mice) |
Starting from 5 days before immunization
(oral) |
50, 100, 150 mg/kg/d for 35 days |
↓ free radical (DPPH) |
↓ Th1, Th17 cells.
↓ p-STAT1, p-STAT3, p-STAT4.
↓ IL-17, IL-1α, IL-1β, TNF-α, CCL1, CCL2, CXCL1, CXCL10, CXCL11. |
| Crocus sativus L. |
EAE
(male C57BL/6 mice) |
Starting from immunization day
(oral) |
500 mg/kg/d
for 21 days |
↑ TAC (FRAP).
↓ NO |
↓ inflammatory infiltration |
| Moringa oleifera. |
CPZ
(Wistar rats) |
Starting from cuprizone diet fed (oral) |
1.875 mg/mL/mouse/d
for 35 days |
↓ NO
↑ CAT, SOD |
↑ neuronal integrity. |
| Olive oil |
EAE
(Dark Agouti rats) |
starting at 11th day
post immunization
(oral) |
representing 10% of calorie intake in the total standard diet for 54 days |
↑ GPx.
↓ LPO, NO. |
↓ NF-κBp65, TNF-α.
↓ LPS, LBP. |
| Copaiba oil |
EAE
(female C57BL/6 mice) |
Splenocytes were obtained from EAE mice at day 20 post immunization
(in vitro) |
100, 50 and 25 µg/mL
for 24 h |
↓ H2O2, NO. |
↓ TNF-α, INF-γ, IL-17 |
| Sesame oil |
EAE
(male C57BL/6 mice) |
Starting from day 3 before the immunization
(oral) |
4 mL/kg/d
for 28 days |
↓ NO
↑ TAC (FRAP). |
↓ inflammatory infiltration. |
Hypericum
perforatum L. extract |
EAE
(C57BL/6 mice) |
Starting from immunization day
(oral) |
18−21 g/kg/d
for 42 days |
↑ TSA.
↓ TOS, OSI. |
↑ MOG, MBP.
↓ inflammatory infiltration. |
Oenothera
biennis L. extract |
EAE
(C57BL/6 mice) |
Starting from immunization day
(oral) |
18−21 g/kg/d
for 42 days |
↑ TSA.
↓ TOS, OSI. |
↑ MOG, MBP.
↓ inflammatory infiltration. |
2.1. Natural Phenolic Compounds
Natural phenolic compounds are plant-derived antioxidants. Phenols, the important secondary metabolites in plants, are mainly produced by the metabolic pathways of shikimic acid [
132]. Phenols are widely found in the roots, stems, leaves, flowers, fruit pulp and seeds of various higher plants. Among the major bioactive substances of plants, phenolic compounds are the most diverse and widely studied class of chemicals. Structurally, all phenolic compounds contain at least one benzene ring and one hydroxyl group. The strong antioxidant capacity of natural phenols is mainly attributed to the hydroxyl group, which can effectively quench free radicals, chelate metal ions, induce antioxidant enzyme activity and regulate the in vivo antioxidant signalling pathways [
133]. Natural phenolic compounds can be classified into flavonoids and non-flavonoids depending on their chemical structures.
2.1.1. Flavonoids
Anthocyanin (ANT), a flavonoid, is one of the polyphenolic compounds with the highest concentration in food. Natural ANT, widely found in angiosperms, are water-soluble natural pigments that give petals and fruits purple, red and blue colors. ANT have anti-inflammatory, anti-tumour, anti-ageing, and metabolism-regulating effects, which are related to their excellent antioxidant capacity [
134]. The antioxidant mechanisms of ANT mainly include the reduction of ROS accumulation and scavenging of free radicals, activation of the enzymatic antioxidant system and attenuation of DNA damage. Oral administration of ANT to rats with EB-induced MS was found to reduce demyelination, decrease the levels of HNE-histidine (HNE-His) and MDA and increase the activities of Non-protein sulfhydryl group (NPSH), GSH and SOD. Administration of ANT also resulted in near-normal levels of protein carbonylation, NOX and inflammatory cytokines in rats with EB-induced demyelination. In addition, EB increased MPO activity, and MPO expression was associated with inflammatory cell infiltration. The presence of neutrophils and the release of active substances can lead to the increase of oxidative stress, the decrease of ion pump activity, and the death of oligodendrocytes. ANT treatment partially inhibited the aggravation of interleukin-induced inflammation and decreased EB induced ion pump activity. Treated with ANT also reduced MPO and inflammatory cell infiltration in rat brain tissue, thus preventing the harmful effect of EB on demyelination [
135].
Hesperidin (HP), a natural flavonoid widely found in the plant kingdom (mainly in citrus plants), is the main pharmacological component of the fruits of the genus
Citrus, family Rutaceae, with various biological activities including anti-inflammatory, antioxidant, and tumour metabolism regulatory functions [
136]. EAE-induced lipid peroxidation caused a significant increase in TBARS levels, leading to irreversible CNS injury. In addition, EAE inhibits the enzyme (SOD, CAT, and GPx) and non-enzyme (GSH) antioxidant defense systems in brain tissue. Oxidative stress plays an essential role in MS because it can lead to significant cell death and neuronal damage in the brain. In this case, the effect of EAE on the brain may mainly depend on the increase of oxidative stress. Therefore, antioxidants such as HP may be helpful for EAE-induced demyelination. Intraperitoneal injection of HP after the onset of EAE reduced the disease severity by decreasing the levels of interleukin (IL)-17, IL-1β and tumor necrosis factor (TNF)-α, inhibiting lipid peroxidation (TBARS) and increasing the antioxidant markers (GSH, CAT, GPx and SOD) in brain tissues, thereby reducing histopathological damage in the brain [
137].
Licochalcone A is a flavonoid extracted from liquorice roots and is a species-specific component of
Glycyrrhiza inflata. Licochalcone A is the most abundant among the chalcones isolated and identified in liquorice. Compared with the normal control group, spleen cells from the EAE group showed high levels of oxygen free radicals (H
2O
2, NO) and cytokines (TNF-α, IFN- γ, and IL-17). As observed in MS and EAE, long-term exposure to oxidative stress environment will destroy the biological structure and lead to significant cell destruction, causing direct damage to the myelin sheath. It has been reported that IFN- γ (Th1 cytokine), IL-17 (Th17 cytokine), and TNF- α play a vital role in the induction and severity of MS/EAE. In the acute phase of EAE, the expression level of IFN- γ and TNF- α was parallel to the severity of clinical symptoms and the degree of inflammatory cell infiltration. Licochalcone A can alleviate the rise in clinical scores and weight loss involved in EAE and terminates the excessive immune response, which is attributed to its property of inhibiting the levels of interferon (IFN)-γ, TNF-α and IL-17. In addition, it can reduce the expression of the oxidative markers H
2O
2 and NO, as confirmed by both in vitro and in vivo experiments [
138].
Flavonoids in
Scutellaria baicalensis and
Acacia catechu exert a variety of therapeutic effects, including anti-inflammatory, antiviral, antibacterial, and anticancer activities. Unlike traditional natural flavonoids, flavocoxid (FVC) is a purified mixture containing baicalin and catechin that acts as a dual inhibitor of cyclooxygenase (COX)-2 and 5-lipoxygenase (LOX) [
139]. Th1/Th17 cells play an essential role in EAE. These cells migrate from the spleen to the CNS. They are re-stimulated by perivascular antigen-presenting cells (APCs) and promote microglia activation by releasing pro-inflammatory cytokines. Activated microglia have two different phenotypes, the classical M1 and the alternatively activated M2. M1 cells express iNOS, produce high levels of oxidative metabolites and pro-inflammatory cytokines, and destroy myelin sheath. M2 phenotype has neuroprotective and anti-oxidative effects. In addition to its anti-inflammatory properties, FVC serves as an antioxidant to reduce ROS, including •OH, O
2•
− and H
2O
2. FVC was shown to attenuate the severity of EAE, reduce the levels of COX-2, 5-LOX, iNOS, IL-12, IL-23, IL-17 and IFN-γ and increase the levels of IL-10 in the spinal cord of mice. In addition, oral administration of FVC decreased the M1 phenotypes and increased the proportion of M2 phenotypes in macrophages and microglia. Furthermore, FVC inhibited CD4
+ T cell Th1/Th17 differentiation [
140].
Luteolin was originally isolated from the stems and leaves of the mignonette plant. Recent studies have revealed that luteolin is predominantly present as glycosides in a variety of herbs, vegetables and fruits, such as
honeysuckle,
Schizonepeta and pomegranate. Luteolin has multiple pharmacological properties such as antibacterial and antiviral properties. In addition, it has exhibited excellent safety and efficacy in a variety of pre-clinical models as an anti-inflammatory, antioxidant and neuroprotective agent [
141]. Furthermore, luteolin is a viable therapeutic agent for oxidative stress associated with autoimmune diseases. NF-κB activation in MS patients leads to extreme expression of chemokines and cytokines, macrophage inflammatory protein-1α (MIP-1α), one of the most frequently expressed chemokines throughout CNS inflammation, involved in MS/EAE progress. Neuronal and oligodendrocyte apoptosis is considered an important feature of MS, which may be attributed to oxidative stress-induced cell membrane damage and impaired myelination, and activation of caspase-3 within neurons leads to axonal degenerative changes. cAMP has shown a critical role in neuronal growth, differentiation and survival. cAMP levels were decreased in the cerebrospinal fluids of MS patients, and inhibition of the cAMP/CREB signaling axis attenuated CNTF expression. Compared to EAE mice, luteolin-treated EAE mice demonstrated improved clinical motor function, reduced inflammatory infiltration of brain tissues, upregulated ciliary neurotrophic factor (CNTF) expression, significantly increased levels of cAMP and total antioxidant capacity (TAC) of the brain and significantly reduced levels of cleaved caspase 3, nuclear factor kappa-B (NF-κB) and macrophage inflammatory protein (MIP)-1α [
142].
Epimedium flavonoids (EF) are a total flavonoid component extracted from the stem and leaves of plants belonging to
Epimedium spp. of the family Berberidaceae, containing mainly icariin and icariside. EF exhibited good neuroprotective effects in animal models of Alzheimer’s disease, traumatic brain injury, ischaemic stroke and depression. Microglia and astrocytes were activated in the brain and spinal cord of EAE model, resulting in the production and release of pro-inflammatory molecules (IL-1β, TNF-α, NO) and the upregulation of iNOS activity. NF-κB is a family of transcription factors critical for immunity and inflammation. It regulates many biological processes, including sensitivity to inflammatory cytokines, oxidative stress, and apoptosis. NF-κB-dependent CNS immune cell activation and oligodendrocyte death are essential pathophysiological features of multiple sclerosis. Oral administration of EF to rats with EAE resulted in the attenuation of neurological deficits, demyelination and inflammatory infiltration, activation of astrocytes in the spinal cord, and inhibition of IL-1β, TNF-α and NF-κB expression. In addition, it resulted in upregulated level of nerve growth factor (NGF) and improved the ultrastructure of myelin and axons. Furthermore, oxidative stress-related iNOS and NO in the spinal cord of rats with EAE were downregulated after treatment with EF [
143].
Kolaviron (KV), derived from traditional African medicine, is a bioflavonoid isolated from the seeds of Garcinia kola. Previous in vitro and in vivo studies have revealed that KV has many pharmacological properties such as anti-inflammatory, antibacterial, anti-apoptotic and antioxidant properties [
144]. CPZ significantly downregulated SOD levels within the rat prefrontal cortex, exacerbated lipid peroxidation, reduced spatial memory and disrupted the integrity of neuronal and non-neuronal cells. Moreover, during CPZ intervention, GSH content decreased, and excessive ROS/RNS were produced to induce CNS damage. CPZ poisoning can lead to the formation of giant mitochondria and oxidative stress, resulting in the interruption and shortage of energy flow. The abnormal energy flow impairs the function of the endoplasmic reticulum, causes extensive proliferation of the endoplasmic reticulum and decreases hydrophobin synthesis, aggravating the disintegration of oligodendrocytes and myelin sheath. However, these changes were significantly reversed in KV-treated rats. Such anti-demyelinating pharmacological effects of KV are mediated by its inherent antioxidant properties to counteract CPZ-induced oxidative damage. Therefore, KV, a natural compound with neuroprotective effects, deserves further study as a potential anti-MS therapeutic agent [
145].
Quercetin (Que), a polyhydroxy flavonoid, is a secondary plant metabolite that is widely found in a variety of vegetables and fruits such as hawthorn, broccoli and onions, as well as in herbs such as
Forsythiae Fructus,
Gynostemma pentaphyllum and
Houttuynia cordata. Que exhibits prominent antioxidant, free radical scavenging, anti-inflammatory and immunomodulatory properties [
146]. The researchers found that the demyelination of pons caused the decrease of Na
+, K
+-ATPase activity, directly damaging axonal pulses’ transmission. The measurement of blood biomarkers may be one of the crucial methods to evaluate the oxidative status and disease severity of neuroinflammation. In some CNS inflammatory diseases, oxidative stress and inflammation interact with redox dysfunction in erythrocytes. Erythrocytes in MS patients are more prone to cytolysis, and erythrocyte membrane fluidity defects are closely related to the intensity of demyelination and the process of lipid peroxidation. In rats with EB-induced demyelination, treatment with Que increased Na
+ and K
+ ATPase activity in the pons and cerebellum and inhibited the EB-induced increase in acetylcholinesterase activity in whole blood and lymphocytes. EB decreased the activity of CAT in whole blood and increased serum MDA levels in rats. However, Que reversed this change, significantly increased the antioxidant capacity and alleviated the EB-induced inflammatory cell infiltration and behavioural deficits in rats [
147].
Phloretin (Phl) is a dihydrochalcone compound found in common fruits such as apples and strawberries. Phl has immunomodulatory properties and has been widely studied for its natural antioxidant properties. In vivo studies have revealed that Phl alleviated the clinical signs and neuroinflammation in mice with EAE. Nrf2 is a significant regulator of antioxidant response. Many studies have determined that its activation can drive macrophages to produce anti-inflammatory phenotypes and reduce neuroinflammation and neurodegeneration in CNS diseases. Nrf2 activation is mediated by AMPK activation, AMPK plays a crucial role in restoring cellular energy homeostasis. It has been found that Phl stimulates autophagy in an AMPK-dependent manner and activates the Nrf2 pathway through autophagy-mediated degradation of Keap1 to reduce macrophage-derived NO and ROS, ultimately driving macrophage polarisation toward a protective phenotype. These findings suggest that Phl may be a potential therapeutic agent for inflammatory demyelinating diseases of the CNS [
148].
2.2. Terpenoids
The plant constituents formed by the derivation of mevalonate are called terpenoids. Terpenoids are the most numerous and diverse natural products, which can be classified into monoterpenes, sesquiterpenes, diterpenes, triterpenes, and polyterpenes according to their chemical structures. These compounds have numerous biological activities, including anti-inflammatory, antioxidant and immunomodulatory, and are widely used in various diseases, which have attracted increasing attention in recent years [
161].
Glycyrrhizae Radix et Rhizoma is a commonly used herb in traditional Chinese medicine. 18β-glycyrrhetinic acid (GA) is a pentacyclic triterpene derivative isolated from
Glycyrrhizae Radix et Rhizoma, which is a hydrolyzed metabolite of glycyrrhizinic acid and liquiritigenin. Due to its antioxidant, anti-inflammatory and anti-cancer effects, GA is often used to treat digestive and neurological disorders. As we all know, inflammation is significant in autoimmune diseases such as MS. The excessive production of pro-inflammatory cytokines and chemokines will cause the death of nerve cells and lipid membrane disorder in the CNS of EAE mice, which lead to the generation of reactive oxygen species, aggravating the oxidative damage of neurons and glial cells. EAE induced a significant increase in oxidative stress and lipid peroxidation (elevated TBAR levels, decreased GPx, SOD, CAT and GSH levels) in the mouse brain. In addition, EAE mice had many inflammatory cells infiltrating and highly expressed pro-inflammatory factors and apoptosis markers (IL-17, TNF-
α and IL-1
β, Caspase-3). In contrast, GA treatment significantly reversed the oxidative histological and pathological changes of EAE [
162].
β-Caryophyllene (BCP), a natural sesquiterpene found in many plants, is a potent anti-inflammatory compound. In MS, when microglia and macrophages are activated, they release many cytotoxic mediators, including IFN-γ, TNF-α, NO, and ROS, which may cause CNS tissue damage. IFN-γ is involved in macrophage activation, T cell differentiation, and regulation of T lymphocyte function, which is particularly critical in autoimmune diseases. TNF-α is an important factor causing neuronal damage, and its production is regulated by NF-κB. Animal studies revealed that BCP treatment reduced the clinical severity and pathological damage of EAE and inhibited the production of TNF-
α, IFN-
γ and IL-17 as well as oxidative markers H
2O
2, and NO in CNS. Consistent with the in vivo results, in vitro intervention with BCP in splenocytes of EAE mice significantly reduced oxygen free radicals and inflammation [
163].
Ginkgolide K (GK) is a diterpene lactone compound extracted from the leaves and roots of Ginkgo, which has antagonistic properties against platelet-activating factor receptors. Evidence suggests that Ginkgo extracts have neuroprotective effects under hypoxia/ischemia, Alzheimer’s disease, and anxiety [
164]. Intraperitoneal injection of GK improved behavioral dysfunction and demyelination in CPZ mice. The nuclear factor Nrf2 mainly modulates the endogenous cellular antioxidant response. Activation of Nrf2 suppresses oxidative stress and inflammatory responses in the CNS in MS. The Nrf2 agonist dimethyl fumarate has been approved for clinical treatment of MS. Astrocytes are the primary cell type that activates Nrf2 under pathological conditions in the CNS. Nrf2 activation is regulated by the IGF/PI3K/Nrf2 signaling pathway. Studies have found that IGF-1 knockout mice have reduced Nrf2 expression and increased oxidative stress and apoptosis. IGF-1 also reduces neuronal death by activating the PI3K/Nrf2 pathway. GK treatment upregulated Nrf2/HO-1 and downregulated p-NF-κB/p65 in astrocytes, inhibiting oxidative stress-related NO and iNOS in astrocytes. In addition, activation of Nrf2 after GK treatment modulated the IGF/PI3K signaling pathway to attenuate oligodendrocyte apoptosis in corpus callosum [
165].
Bixin is isolated from the seeds of
Bixa orellana and approved by FDA for use in the food and cosmetic industries, which can cross the blood–brain barrier and can prevent oxidative DNA damage and inhibit lipid peroxidation [
166]. Bixin can significantly improve symptoms and inflammatory demyelination damage in EAE mice, and reduce the release of TNF-α, IL-6, IL-8, IL-17, and IFN-γ. The TXNIP/NLRP3 inflammasome is critical in the pathogenesis of CNS disease. The TXNIP/NLRP3 inflammasome is highly expressed in the spinal cord of EAE mice and worsens demyelination. Oxidative stress is one of the major causes of CNS dysfunction in MS, and CD4+ T cells produce large amounts of ROS, which are major mediators of oxidative stress and initiators of the TXNIP/NLRP3 inflammasome. Bixin can clean excess ROS and reduce the number of Th1/Th17 cells, which was achieved by activating Nrf2 signaling pathway and suppressing the activation of the TXNIP/NLRP3 inflammasome [
167].
Olibanum is a hard gel-like resin exuded from
Boszvellia carterii Birdw. and
Boswellia bhaw-dajiana Birdw. Olibanum contains a series of complex components, of which the major active ingredient is 11-keto-beta-boswellic acid (AKBA), a resinous pentacyclic triterpenoid. AKBA has various physiological effects, including anti-infective, antitumor and antioxidant effects [
168]. Treatment with AKBA alleviated demyelination and clinical symptoms in EAE mice. There is evidence that IL-6 amplifies neuroinflammation by activating immune cells of resident and peripheral origin. In MS/EAE, high IL-6 expression in the CNS was associated with elevated p-NF-κB levels. In EAE mice, oxidative damage markers such as lipid peroxides were elevated. iNOS participates in the MS neuroinflammation process by producing oxidants, and the production of iNOS is regulated by the transcription factor NF-κB. Activation of NF-κB leads to upregulation of IL-6 and iNOS-mediated oxidative stress, which is accompanied by downregulation of Nrf2 in a mouse model of EAE. AKBA treatment inhibited NF-κB signal transduction and activated the Nrf pathway, thus correcting the overexpression of IL-6 and iNOS, and alleviating the immune-inflammatory response in EAE. Therefore, AKBA may have a good therapeutic effect on relapsing-remitting MS [
169].
Ginsenosides are triterpene saponins (terpene glycosides), of which ginsenoside Rg
3 is a natural steroid saponin found in high contents in Korean red ginseng. Compared with other ginsenosides, ginsenoside Rg
3 has a wide range of pharmacological activities, and it shows neuroprotective effects in various CNS disease models [
170]. Ginsenoside Rg
3 decreased demyelination and increased the integrity of the blood–brain barrier in EAE mice with CNS. COX-2, iNOS, IL-1β, IL-6 and TNF-α have downregulated after ginsenoside Rg
3 treatment, while Arg-1 and IL-10 levels were elevated. Activated NOX2 and NOX4 can produce ROS released into the intracellular or extracellular space, aggravate the oxidative damage of oligodendrocytes, and lead to progressive demyelination. Ginsenoside Rg
3 could also inhibit the expression of oxidative stress markers, including mtROS, NADPH, NOX2, NOX4 and 4-Hydroxynonenal (4-HNE) in the spinal cord of EAE mice [
171].
Astragaloside (AST) is a tetracyclic triterpenoid saponin from Astragali Radix that has been shown to have good therapeutic effects in CNS demyelinating disorders [
172]. AST administration significantly inhibited severity and blood–brain barrier leakage in EAE mice, reduced inflammatory cell infiltration and ROS production in CNS, upregulated SOD activity, and reduced neuroinflammation by inhibiting iNOS. In addition, different subtypes of T cells affect the occurrence and development of MS. Transcription factors T-bet, RORγt and Foxp3 contribute to the differentiation of CD4 naive T cells into distinct subpopulations. In the presence of IL-12 and T-bet expression, naive T cells can be induced to differentiate into Th1 cells; when TGF-β, IL-6 and retinoic acid receptor related to orphan receptor gamma-t (RORγt) are present together, Th17 cells will be generated. Foxp3 is essential for the development and function of Tregs. AST increased the mRNA expression of transcription factors T-bet and Foxp3 in the peripheral immune system of EAE mice but decreased the expression of ROR
γt to regulate T cell differentiation [
173].
2.3. Phenylpropanoids
Nordihydroguaiaretic acid (NDGA), a phenolic lignan originally isolated from the leaves of
Larrea tridentata, acts as one of the inhibitors of LOX and can suppress the inflammatory reaction. NDGA exerts neuroprotective effects by scavenging ROS and activating HO-1 in EAE mice. The p38MAPK signaling pathway regulates IL-17 synthesis in CD4+ T cells. SGK1 is one of the substrates of p38MAPK, and oxidative stress stimulates SGK expression through a p38/MAPK-dependent pathway. There is a strong correlation between oxidative stress and the p38MAPK-SGK1 pathway in T cell-mediated autoimmune inflammation. The p38MAPK-SGK1 pathway is the confluence of CNS oxidative stress and Th17 inflammatory responses. Oxidative stress activates the p38MAPK-SGK1 pathway and enhances the Th17 phenotype, thereby inducing pathological damage and neurological dysfunction. Oxidative stress p38MAPK-SGK1 pathway may be a central link in EAE, and MS. NDGA attenuates immune inflammation and oxidative stress in EAE mice by inhibiting p38MAPK-SGK1 pathway. Behavioral and pathological myelin damage is improved in NDGA-treated EAE mice, IL-17 and MDA are decreased in the spinal cord, and the antioxidant HO-1 is upregulated [
174].
Caffeic acid phenethyl ester (CAPE) is a natural active compound derived from propolis, a catechol-containing phenylpropanoid derivative with multiple biological activities [
175]. CAPE has significant advantages in antibacterial, antioxidant, anti-inflammatory and anti-cytotoxic properties. CAPE treatment significantly improved the clinical symptoms in EAE rats. In addition, CAPE may reduce myelin damage and destruction by inhibiting NF-
κB activation-mediated inflammatory responses, reducing ROS production, and exerting its antioxidant effects by downregulating iNOS, MDA, MPO, NO levels and increasing GPx, SOD activity in CNS tissues [
176]. It has been found that ROS production is critical for NF-κB activation. However, CAPE inhibits NF-κB activation by inhibiting the interaction of NF-κB protein with DNA, not by maintaining IκB-α expression. CAPE reduces NF-κB activation by reducing the release of reactive oxygen species intermediates in the CNS of EAE rats. Xanthine oxidase (XO) is a physiological source of superoxide anion in eukaryotic cells. Tissue XO activity was significantly increased in the EAE group, while CAPE treatment reduced its expression, alleviating the overgenerated free radicals in the spinal cord of EAE rats.
P-Coumaric acid (p-CA), also known as 4-hydroxycinnamic acid, is a derivative of cinnamic acid and is present in free form in many plants [
177]. Its activity in reducing oxidative stress and inflammatory responses have been demonstrated in various animal models, and it has been extensively studied due to its beneficial effects on CNS disease. Matrix metalloproteinase 9 (MMP-9) is secreted by activated lymphocytes, endothelial cells, and leads to disruption of the blood–brain barrier. It is essential to promote T cell migration to the CNS and induce demyelination during MS. Studies have shown that the expression of MMP-9 is regulated by oxidative stress level. In a CPZ-induced MS mouse model, treatment with p-CA by gavage significantly inhibited high expression of MMP-9 in the brain, protected integrity of the blood–brain barrier, and reduced the levels of the free radical indicator DPPH [
178].
Magnolol (MAG), a phenylpropanoid derived from
Magnolia officinalis, is a hydroxylated biphenyl natural compound. MAG can significantly reverse the clinical symptoms, pain parameters and CNS pathological changes of EAE and reduce the level of cytokines in a dose-dependent manner. Low expression of Nrf2 is associated with oxidative stress-mediated oligodendrocyte apoptosis in EAE. In vitro and in vivo studies have shown that Nrf2 attenuates CNS tissue damage. In contrast, failure of Nrf2 signaling renders the CNS more vulnerable and leads to excessive protein accumulation and neurodegeneration. In MS/EAE, Caspase-3 is a significant player in neuronal apoptosis, and inhibition of Caspase-3 activation in brain tissue can effectively protect neurons and oligodendrocytes. MAG treatment significantly inhibited oxidative stress by downregulating the activities of MDA, NO and MPO, and increased the levels of antioxidants, such as GSH, GST, cat and SOD. In addition, MAG significantly enhanced the antioxidant defense system by increasing the expression level of Nrf2 while decreasing the expression of iNOS and cleaved-caspase-3 in the brain to protect myelin integrity [
179].
Daphnetin (DAP) is a 7, 8-dihydroxycoumarin extracted from
Daphne odora. It has been shown in several preclinical animal studies and cellular experiments to be a potent neuroprotectant that effectively blocks neuronal damage in the CNS [
180]. DAP-treated EAE mice have decreased brain levels of cytokines, including IL-17, INF-γ, IL-6, IL-12a and IL-23a. Overactivated Th1 and Th17 cells and inflammatory responses may induce oxidative stress in the progression of MS/EAE. HO-1 is a stress-responsive enzyme with potent antioxidant activity, and up-regulation of HO-1 expression inhibits inflammation-induced oxidative stress. Studies have shown that knockdown of HO-1 significantly increases the expression of IL-6, TNF-α, and IL-1β. These cytokines may be associated with the generation of Th1 and Th17 cells and the exacerbation of inflammatory pathology in EAE. DAP inhibited the production of IL-1β, IL-6 and TNF-α in LPS-stimulated BV2 cells. HO-1, a typical antioxidant and anti-inflammatory factor, was induced in large amounts in BV2 cells after daphnetin treatment. In addition, a significant elevation of HO-1 was observed in the brains of daphnetin-treated EAE mice, along with downregulation of MDA levels [
181].
2.4. Alkaloids
Piperine, an alkaloid compound belonging to the amide group isolated from the fruit of black pepper (
Piper nigrum), has important pharmacological activities, including in MS/EAE treatment. We all know that iNOS is one of the major oxidation-related enzymes in MS. It catabolizes arginine to generate NO in the environment of CNS injury, which affects the process of cytotoxicity, inflammation, and oxidative stress injury. The high expression of iNOS in the brain hinders the recovery of MS. Nrf2 regulates the expression of numerous antioxidant genes, including HO-1. HO-1 catalyzes the degradation of heme and produces carbon monoxide, ferrous iron, and biliverdin, thereby reducing oxidative damage. Administration of piperine attenuated neurological deficits, inhibited disease progression, reduced demyelination, inflammation, immune cell infiltration, microglia and astrocyte activation, and enhanced the expression of IL-10, Nrf2, HO-1, MBP and NeuN in EAE rats. Piperine also enhanced the total antioxidant capacity (FRAP) and reduced the levels of oxidative stress markers (MDA) in the CNS of EAE rats, while replenishing deficient BDNF and reducing neuronal apoptosis. Similarly, the results were consistent with the EAE model in LPC-induced demyelinated rats treated with piperine [
182,
183].
Matrine (MAT), a tetracyclo quinolizine alkaloid, is the main active ingredient extracted from the dry root of
Sophora flavescens and used in a variety of CNS and peripheral inflammatory diseases [
184]. Intraperitoneal MAT injection effectively inhibited EAE progression and significantly reduced oligodendrocyte apoptosis, microglial activation, and inflammatory factor secretion. The energy and redox state of mitochondria play a decisive role in cellular homeostasis. Mitochondrial dysfunction is associated with the degeneration of myelin sheaths and axons. ROS mainly originates from the electron leakage of mitochondrial complexes. During apoptosis, the abnormal structure and function of mitochondria lead to swelling of organelles, and then the cytoplasm releases Cyt-c and other apoptotic factors. Mitochondrial autophagy, a major pathway for maintaining mitochondrial function, has profound effects on the repair of damaged oligodendrocytes. MAT treatment also reduced the levels of Cyt-c and the oxidative stress marker MDA in CNS tissues. In contrast, autophagy-related proteins Beclin1, LC3 and GPx were upregulated after MAT treatment, thereby enhancing mitochondrial autophagy and alleviating the imbalance of oxidative/antioxidant systems induced by mitochondrial injury [
185].
4.5. Quinones
Shikonin is a class of natural naphthoquinones extracted from the roots of Lithospermum erythrorhizon. Shikonin treatment of EAE mice significantly reduced the extent of corpus callosum demyelination. The level of ROS in MS increased, and the antioxidant defense system in vivo was damaged, which led to the enhanced blood–brain barrier permeability. Various inflammatory factors enter the brain and spinal cord in large quantities. Among them, TNF-α directly induces oligodendrocyte death and oligodendrocyte progenitor cell loss. IFN-γ can promote the inflammatory response of EAE and MS. The accumulation of these cytokines significantly increased the expression level of apoptosis-related proteins in CNS. Conversely, TGF-β is an anti-inflammatory cytokine, mainly produced by T cells, monocytes, astrocytes, and microglia, which can prevent autoimmune response and inflammatory damage. The genes expression of TNF-α, IFN-γ and Bax was enhanced and TGF-β and Bcl2 were reduced in the brain tissue of EAE mice, and shikonin treatment significantly reduced the expression levels of TNF-α, IFN-γ and Bax. In addition, the expression levels of TGF-β and Bcl2 and the activity of GPx1 were significantly increased after shikonin treatment. These results suggest that shikonin has good immunomodulatory and antioxidant effects in EAE and may contribute to the remission of EAE [186].
Thymoquinone (TQ), a benzoquinone, is a natural antioxidant isolated from the seeds of Nigella sativa and present in other plants [187]. Increased IL-17 and IL-17R have been reported in an EAE-induced demyelination mouse model, and treatment with TQ reduced this abnormal expression. In addition, TQ inhibited the development of acute and chronic recurrent EAE in mice, reduced the number of perivascular inflammatory cell infiltrates and increased GSH in the spinal cord, suggesting that TQ may reduce myelin damage in EAE mice by inhibiting oxidative stress [188,189].
4.6. Steroids
Withametelin (WMT), a natural sterol lactone derived from the leaves of Datura stramonium, has been shown to relieve depression and neuropathic pain significantly [190]. Dysfunction of the blood–brain barrier (BBB) plays a critical role in the pathogenesis of MS, with the migration of pro-inflammatory cells and toxic molecules to the brain through the damaged BBB, leading to demyelination and neuronal death. Fourier transform infrared spectroscopy revealed that EAE induced significant changes in myelin biomolecular composition, including protein oxidative damage, lipid peroxidation, increased nucleic acid/carbonyl content, and decreased lipid/protein content. It is well known that Nrf2/Keap-1-mediated oxidative stress and neuroinflammation contribute to neuronal degeneration in EAE models of MS. Oxidative stress in the brain, spinal cord, and optic nerve may cause permanent cellular damage due to the oxidation of cellular components. In MS, low levels of antioxidant enzymes and high levels of reactive oxygen species aggravate CNS damage. WMT treatment significantly attenuated EAE-induced weight loss, neuropathic pain, and motor dysfunction reduced elevated circulating leukocytes and blood–brain barrier disruption, and reversed histopathological changes in the brain, spinal cord, and optic nerve. WMT enhanced the antioxidant defense mechanism and decreased the expression of Keap-1 and iNOS by increasing the expression levels of Nrf2 and HO-1 in the CNS [191].
Guggulsterone (GST) is a natural phytosterol from the resin of Commiphora mukul, which is widely used in preclinical animal studies of CNS diseases such as ischemic stroke, dementia, depression, and autism [192]. PPAR-γ plays an important role in neuroinflammation, and up-regulation of PPAR-γ can reduce pathological expression in EAE models. Elevated PPAR-γ enhances remyelination in MS by suppressing T cells. It has been reported that PPAR-γ agonists reduce ROS production, protect mitochondria, and promote oligodendrocyte differentiation and maturation. Conversely, inhibition of JAK/STAT-mediated glial activation was neuroprotective, reduced interleukin and Th1 cell differentiation, and attenuated oxidative damage. GST improves behavioral deficits (spatial cognitive memory, grip and motor coordination) and increases the expression of the myelin marker MBP in EB demyelinated rats. GST also modulates neurotransmitter levels by increasing acetylcholine, dopamine, serotonin and decreasing glutamate. In addition, GST ameliorates inflammatory cytokines (TNF-α, IL-1β) and oxidative stress markers (AchE, SOD, CAT, MDA, GSH, NO) to prevent EB-induced apoptosis. These effects were associated with the dowregulation of JAK/STAT and upregulation of PPAR-γ signaling pathways by GST [193].
4.7. Other Compounds
Sulforaphane (SFN) is an organosulfur compound derived from cruciferous vegetables (e.g., cauliflower). SFN exerts neuroprotective effects through its antioxidant effect in CNS diseases such as Alzheimer’s disease, Parkinson’s disease, epilepsy, etc. It is reported that ROS reduces the integrity of BBB and facilitates the entry of peripheral immune cells into the CNS. Infiltrated leukocytes produce ROS and induce oligodendrocyte and axon damage. In addition, reactive microglia produce peroxynitrite, the main mediator of oxidative stress and neuronal excitotoxicity, thus driving the neurodegenerative process in MS. Nrf2 is a redox-sensitive transcription factor. Previous studies have shown that Nrf2/ARE transcription pathway is crucial for cell defense against oxidative damage. SFN treatment inhibits inflammatory infiltration, demyelination and upregulation of iNOS and NO in the spinal cord of EAE mice. Another study showed that SFN protected the blood–brain barrier in EAE mice and by upregulating Nrf2/ARE pathway to activate the antioxidant HO-1 and NADPH quinone oxidoreductase 1 (NQO1) expression levels. In addition, SFN treatment can inhibit Th17 response and enhance the release of IL-10. These results suggest that SFN inhibits the development of EAE in mice through its antioxidant and anti-autoimmune inflammatory activities [194,195].
3H-1,2-dithiole-3-thione (D3T) is a compound containing a five-membered cyclic sulfur structure extracted from cruciferous vegetables. D3T effectively induces activation of cellular antioxidant and anti-inflammatory defense systems and provides protection in a variety of disease models. In MS/EAE, antigen-presenting cells activate naive T cells and produce inflammatory cytokines to promote the differentiation of encephalitogenic CD4+ T cells. Dendritic Cells (DCs) have been shown to play a key role in promoting the development of pathogenic Th1/Th17 cells. The proinflammatory cytokines IL-12 and IL-23 produced by DCs have been shown to be critical for Th1 and Th17 differentiation, respectively. The administration of D3T after the onset of EAE effectively prevents disease progression. Pharmacological studies have shown that D3T inhibits dendritic cell activation, suppresses the differentiation of Th1 and Th17, and inhibits microglia activation and inflammatory cytokine expression. In vitro experiments revealed that D3T strongly induced Nrf2 and HO-1 expression and enhanced antioxidant activity in LPS-stimulated dendritic cells [196].
C-Phycocyanin (C-Pc) is a photosynthetic pigment isolated from Spirulina platensis with significant effects in regulating excessive oxidative stress, inflammatory damage and immune responses. There was a massive inflammatory infiltration consisting of lymphocytes and macrophages/activated microglia in animals with EAE. Microglia comprise about 10% of all brain cells and are the first line of CNS immune defense. Most studies have shown that activated microglia exacerbates MS/EAE pathogenesis by producing neurotoxic molecules, proinflammatory cytokines, and oxygen-free radicals. Furthermore, IL-17 and IL-6 are the primary effector cytokines in MS, and their knockout mice are protected from EAE. Therefore, it is necessary to intervene in immune cell-mediated inflammatory responses and oxidative damage. C-Pc can ameliorate clinical deterioration, reduce Inflammatory macrophage/microglia infiltration in spinal cord tissue and IL-6, IL-17 in the brain and serum, and regulates oxidative stress parameters (lower MDA, higher GSH) in the peripheral blood of EAE mice, thereby slowing oxidative damage and increasing myelin repair and regeneration in the CNS [197].
4.8. Plant Extracts
Artemisia dracunculus L. is a perennial herb of the Asteraceae family and is an important herb in traditional medicine in many countries. Modern pharmacological studies have demonstrated that it has inhibitory effects on inflammation and oxidative damage, and hepatoprotective and neuroprotective effects [198]. In MS/EAE, Th17 cells can produce IL-17 and IL-23. IL-17 can inhibit the differentiation and maturation of oligodendrocytes. Moreover, IL-17 can enhance the apoptosis-induced effect of TNF-α in oligodendrocytes, which is mainly related to mitochondrial dysfunction, ROS generation, and cell cycle arrest. Administration of A. dracunculus aqueous extract alleviated weight loss, inflammatory infiltration and demyelination of CNS in EAE mice. After treatment, serum levels of inflammatory cytokines, including IL-17 and IL-23, were reduced, while antioxidant levels (FRAP) were increased [199].
Olive leaves are widely used as a medicinal plant in the Mediterranean region. Some evidence suggests that it contains high levels of phenolics with antioxidant effects to prevent and treat neurodegenerative diseases. The reduction of SIRT1 exacerbates the progression of various neurodegenerative diseases, including MS. Inhibition of SIRT1 expression may contribute to microglial activation and neuroinflammation. SIRT1 has been shown to affect the redox properties of cells and reduce oxidative stress by regulating FOXO3a, resulting in increased activity of CAT and SOD. Based on these facts, SIRT1 is a promising target for MS therapy. Oral administration of olive leaf tea combined with olive leaf extract intraperitoneally in EAE mice attenuated the severity of MS and suppression of SIRT1, upregulated antioxidant enzymes (SOD1, SOD2 and GPx1) and M2 microglia, and inhibited M1 phenotype to maintain myelin integrity [200].
Melilotus Officinalis is an herb from traditional medicine, mainly distributed in East Asia, the Middle East and the eastern shores of Mediterranean, with good anti-inflammatory effects [201]. Disruption of Th1/Th2 balance and oxidative damage and apoptosis of oligodendrocytes play a key role in the pathogenesis of MS. IL-6, TNF-α, IL-12, and IFN-γ are secreted by Th1 cells, but Th2 cells release anti-inflammatory cytokines, such as IL-4, and IL-5, which are vital factors affecting axonal and myelin damage. Prophylactic administration of Melilotus Officinalis extracts attenuated clinical symptoms and pro-inflammatory factors such as IL-6, TNF-α and IFN-γ in the corpus callosum of EAE mice. This herbal extract also promoted the expression of anti-inflammatory cytokines and antioxidant enzymes (CAT, GPx1), thereby maintaining the structural integrity of myelin[202].
Xanthoceras sorbifolia Bunge has long been used in China as traditional folk medicine [203]. Its fruit shell extract (NE) contained high natural phenolic compounds and potent antioxidants. In vivo results showed that oral administration of NE effectively improved clinical disease severity and reduced CNS demyelination in EAE mice. The phosphorylation of STAT1 and STAT3 was highly expressed in CNS of EAE mice, which promoted the polarization of Th1/Th17 cells. NE inhibited Th1 and Th17 cell differentiation via modulating JAK/STAT signaling pathway and reduced the entry of brain inflammatory immune cells into the CNS. In addition, NE exhibited extremely strong antioxidant capacity in vitro and reduced the level of DPPH free radicals [204].
Saffron comes from the flowers of Crocus sativus L., a common medicinal plant whose neuroprotective effects in CNS disorders such as depression, anxiety, Alzheimer’s, Parkinson’s, and epilepsy have received much attention in recent years. In vivo studies showed that oral administration of saffron extracts significantly delayed the onset of EAE disease and attenuated clinical symptoms and CNS inflammatory cell infiltration in mice. Oxygen and nitrogen free radicals lead to lipid peroxidation in MS/EAE, exacerbating neuronal and oligodendrocyte damage. Improving antioxidant enzyme activity is beneficial in preventing free radical-mediated CNS inflammatory response and tissue damage. In addition, total antioxidant capacity (TAC) in the serum of EAE mice treated with saffron extracts was significantly increased [205 ]。
辣木是一种在许多热带和亚热带国家广泛生长的树。辣木的种子、叶子、花和油被广泛用于东南亚传统医学。辣木具有良好的抗氧化、抗糖尿病、抗高血脂、心血管和神经保护作用。CPZ 诱导的脱髓鞘导致大鼠记忆障碍、皮质和海马氧化应激增加(抑制 CAT、SOD、NO 释放增加)和神经元损伤。在多发性硬化症等神经退行性疾病中,硝基酪氨酸阳性蛋白聚集体在细胞内有显着的积累。然而,辣木的管理显着逆转 CPZ 引起的神经病理缺陷和硝化应激,同时增强大鼠大脑的抗氧化能力 [ 206]。橄榄油是从橄榄果实中提取的,在橄榄油中已检测到 200 多种化合物,包括甾醇、类胡萝卜素、三萜醇和酚类。EAE中的微生物群产物(LPS和LBP)与氧化应激呈正相关。LPS和LBP诱导外周免疫系统激活并增加血脑屏障的通透性,导致对CNS的氧化损伤持续存在。用橄榄油灌胃 51 天的 EAE 大鼠大脑、脊髓和血液中的细菌 LPS 和 LPS 结合蛋白 (LBP) 减少。LPO 和 NO(脂质和蛋白质氧化的指标)被下调,抗氧化酶 GSH 和 GPx 的活性增加,从而减缓对中枢神经系统髓鞘的氧化损伤 [ 207 ]。
苦配巴油 (COP) 是一种从苦瓜属植物中提取的油树脂,是南美传统医学中重要的植物药,具有显着的杀菌和抗炎作用。H 2 O 2广泛存在于 MS/EAE 的中枢神经系统中,长时间暴露于高浓度 H 2 O 2会导致不可逆的细胞损伤。与 H 2 O 2类似,NO 对 MS 中的髓鞘损伤负责。NO 具有神经毒性,由巨噬细胞和其他免疫细胞大量产生。体外实验证实,EAE 小鼠脾细胞过度产生氧化介质(H 2 O 2, NO) 和促炎因子 (IFN- γ , TNF - α和 IL-17) 在 MOG 35-55和 ConA 的刺激下, 与 COP 共培养 24 和48 小时 [ 208 ]。
芝麻是许多亚洲国家的传统保健食品,含有许多脂溶性抗氧化剂。免疫后每天对EAE小鼠腹腔注射芝麻油可有效延缓EAE的发作并减轻临床症状。芝麻油能显着提高血清的总抗氧化能力,抑制NO的产生。与芝麻油处理的小鼠相比,在 EAE 小鼠中观察到典型的脑炎性细胞浸润。这一结果表明,芝麻油可有效预防 EAE 的疾病进展,这可能与抑制氧化应激有关 [ 209 ]。
Oenothera biennis L. 属于 Onagraceae 家族,长期以来一直在民间医学中用作良好的天然抗炎和抗氧化剂 [
210 ]。
贯叶连翘属于金丝桃科,其有益效果已在治疗抑郁症、肿瘤和细菌感染方面得到证实 [
211 ]。最近,
O. biennis和
H. perforatum的治疗作用关于 EAE 的报道。巨噬细胞和其他免疫细胞产生的氧自由基和氮自由基参与 MS 的脱髓鞘和轴突损伤。抗氧化剂可以防止自由基引起的脑组织损伤。MS/EAE 脑和脊髓组织增加 TOS 和 OSI 水平并降低 TAS 表达。结果表明,
O. biennis和
H. perforatum提取物降低了 EAE 小鼠大脑的 TOS 和 OSI 水平,增加了 TAS 水平,减少了临床症状和髓鞘损伤。还观察到 EAE 小鼠在血管壁、神经元细胞质和细胞间质空间中有淀粉样蛋白沉积。这些异常表达在
O. biennis和
H. perforatum中显着消除提取物处理组 [
212 ]。