Plant-based foods, such as fruits, contain natural active principles, ranging from primary metabolites (nutritive factors, vitamins, and minerals) to secondary metabolites (phytochemicals). These bioactive principles play significant roles in mitigating several chronic diseases. Some epidemiological studies have identified antioxidant-rich secondary metabolites, such as flavonoids and anthocyanin, for their anti-inflammatory, antiproliferative, and ameliorative roles in neurological disorders. Generally, natural polyphenols express their neuroprotective capacity by relying on their mechanism ability to cross the blood-brain barrier to scavenge the pathological concentrations of reactive oxygen and nitrogen species. Equally, polyphenols modulate a series of mediating cell-signaling pathways of pathological diseases.
- anti-inflammation
- biomechanism
- biomolecules
- fruits
- Musa species
- neurodegenerative diseases
- neuroactivity
1. Overview and Prevalence of Neurodegenerative Diseases
Neurodegenerative diseases essentially relate to any pathological condition that primarily affects the neuron [1]. They affect the central nervous and are typified by the regression and progressive decline of neurological functioning and cognitive deficit [2], which results in major conditions, such as Alzheimer’s, Parkinson’s, and dementia [3][4]. This challenge is particularly devastating in aging populations, with Alzheimer’s disease affecting about 40 million people globally [5][6]. Exposure to multiple factors (environmental and genetic) contributes to the onset of neurodegenerative diseases. Neurotoxic metal pollutants, such as mercury, lead, cadmium, and arsenic, have been identified with Alzheimer’s and Parkinson’s disease, oxidative stress, neuronal death, mitochondrial dysfunction, modulation of metal homeostasis, and aggregation of α-synuclein proteins [7][8][9]. A schematic description of the aforementioned is depicted in Figure 1. It is even more concerning that these environmental factors can cause damage to the neurologic system via epigenetic mechanisms and then trigger neurodegenerative disease in later years [9][10]. Neurodegeneration consists of a series of pathways that have been closely linked to its inflammatory process and, in particular, the pro-inflammatory cytokines implicated in the pathogenesis of functional and neurologic impairment [11].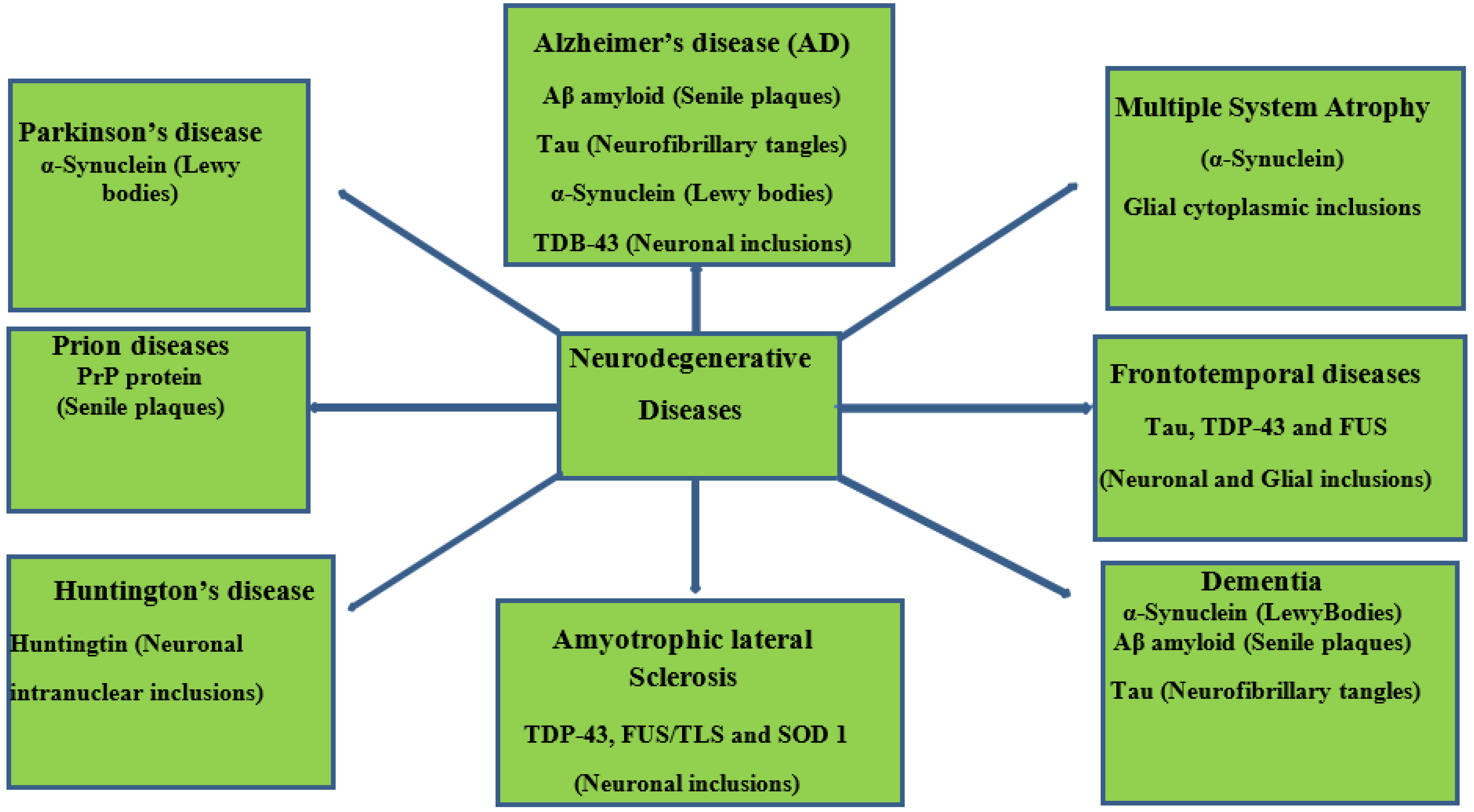
Figure 1. Schematic representation of neurodegenerative diseases, their pathology, and target proteins.
The burden of neurological disorders and conditions has necessitated reliable data to enhance effective health planning approaches. Epidemiological data for neuro conditions, particularly Parkinson’s, dementia, and amyotrophic lateral sclerosis, have been reported in the last two decades [12][13][14][15][16]. Frontotemporal dementia is typically identified in middle age, with reports of about 13% occurrence in people below the age of 50 [17]. Furthermore, Ref. [17] a systematic analysis of a demographics-based study was carried out and estimated about 2–31 frontotemporal dementia incidences per 100,000 people in Europe and developed countries of Asia and North America [17]. An average occurrence of Multiple System Atrophy (MSA) (3 per 100,000 people), average onset age (54–61 years), and demographic prevalence in Europe and North America have been reported in various studies [18][19][20].
2. Fruits in Neurodegenerative Prevention/Management
The brain is especially a centrally significant organ of the nervous system which requires a healthy diet, with fruits potentially offering a wide range of rich nutrient supplies [21]. Fruits represent a major dietary component across Western and Asian demographics. They have been reported to have positive synergy with and relation to chronic disease management [22][23][24][25]. Examples of these common fruits include Malus domestica, Persea americana, Musa sinensis, Citrus limon, Musa paradisiaca, Pyrus communis, Citrus sinensis, Fragaria ananassa, and Ananas comosus (Figure 2).
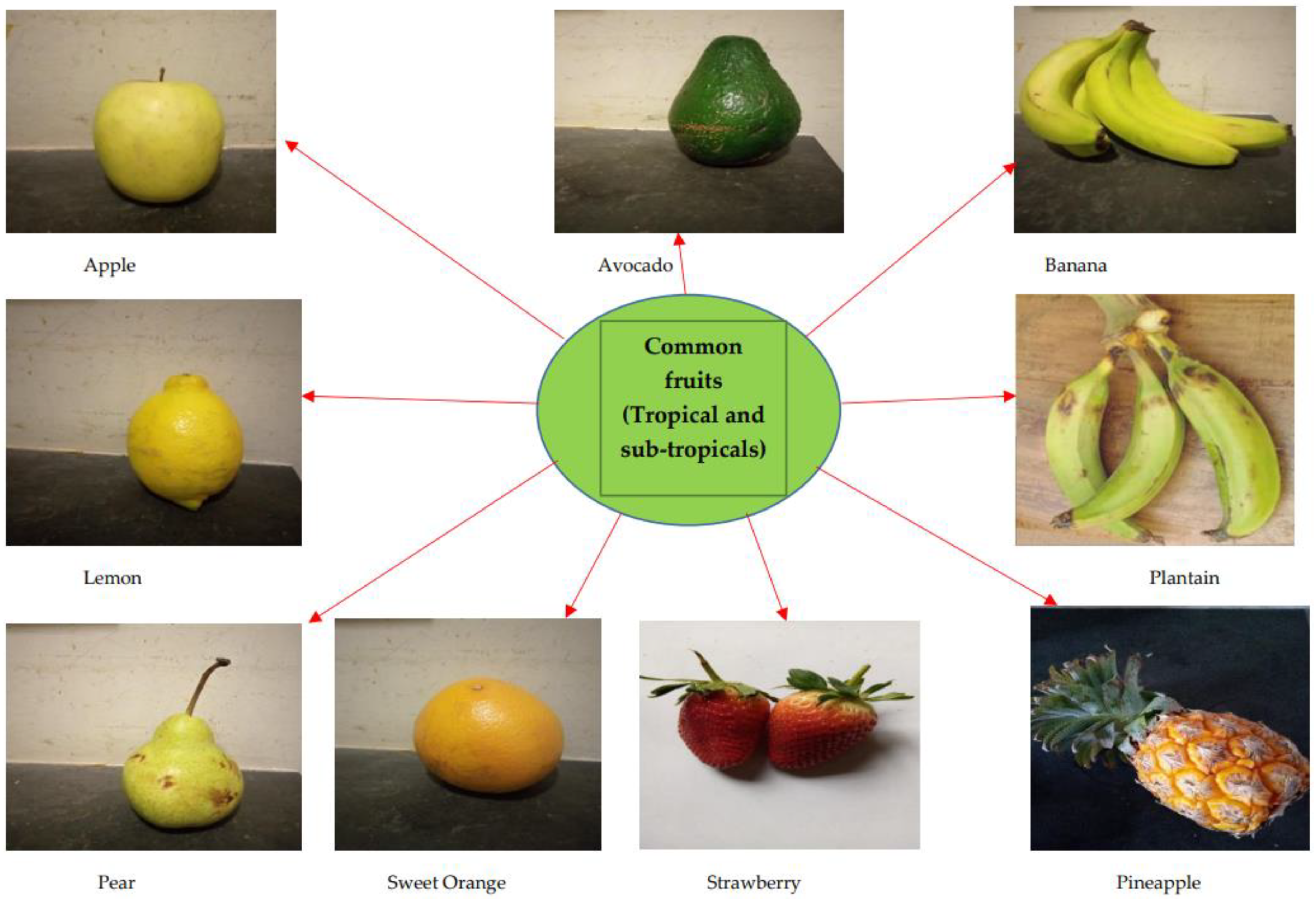
Figure 2. Commonly consumed fruits with bioactive capacities; including Banana and Plantain.
Date palm fruits (Phoenix dactylifera) have been reported in studies for their potential biological capacity in nephroprotective, hepatoprotective, and anticancer activity [26][27][28]. Recently, the inhibitory effect of avocado juice against trypsin aggregation has been reported: a formation process associated with several neurological diseases [29]. Similarly, other reports identified the beneficial role of berries (blackberry and blueberry) with regard to the obstruction of the central nervous system and cognitive deficit [30][31]. The cactus pear (Opuntia ficus-indica) fruit has also been reported to have the biological capacity to modulate neuron excitation in a distributive manner across sectors of the brain [32]. Over time, a host of bioactive molecules cutting across nutritive and antinutritive factors, vitamins, minerals, and secondary metabolites have been screened, evaluated, and elucidated in several species and varieties of banana and plantain fruits, as shown in Table 1.
| Active Molecules (Nutritive Factors and Secondary Metabolites) | Active Molecule Constituents in Banana and Plantain Fruit Components (Pulp and Peel) |
Neuromechanism-Related Protein/Gene Targets |
|---|---|---|
Tannin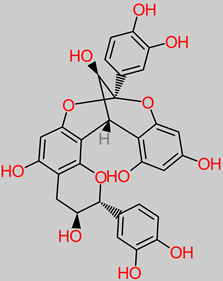 |
|
|
Phenolic acid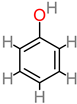 |
|
|
Quercetin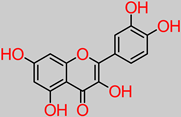 |
|
|
Rutin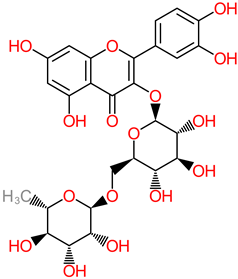 |
|
|
Carbohydrates |
||
Lipids |
||
Magnesium |
|
|
Zinc |
||
Copper |
||
Alkaloid |
|
|
Saponin |
|
|
Phytate (Phytic acid) |
|
|
Vitamins B |
|
|
Anthocyanin |
|
|
Vitamin E (Tocopherol) |
|
|
Selenium |
||
Phytosterol |
|
|
Terpenoids |
|
|
Glycosides |
|
|
Anthraquinone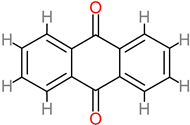 |
|
|
Arginine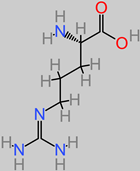 |
|
|
β-carotene |
|
|
Lycopene |
||
β-cryptoxanthin |
|
|
β-sitosterol |
|
|
Sesamin |
- M. sapientum/M. acuminata peel extracts (methanol) [110]. | |
Myricetin |
|
|
Catechin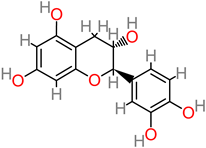 |
|
|
Vitamin C (ascorbic)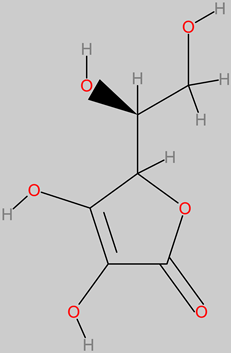 |
|
3. Neuroprotective Mechanistic Narratives of Active Principles and Mineral Elements Listed in Banana and Plantain fruits
3.1. Tannins
3.2. Phenolic Acid
3.3. Quercetin
3.4. Rutin
3.5. Carbohydrates
3.6. Lipids
3.7. Magnesium
3.8. Zinc
3.9. Copper
3.10. Alkaloid
3.11. Saponin
3.12. Phytate
3.13. Vitamin B
3.14. Vitamin C (Ascorbic Acid)
3.15. Vitamin E (Tocopherol)
3.16. Selenium
3.17. Phytosterol
3.18. Terpenoid
3.19. Glycosides
3.20. Anthraquinones
3.21. Anthocyanins
3.22. Arginine
3.23. β-Carotene
3.24. Lycopene
3.25. β-Cryptoxanthin
3.26. β-Sitosterol
3.27. Sesamin
3.28. Myricetin
3.29. Catechin
| Bioactive Compounds | Biological Mechanisms of Action Related to Neuroactivity | Bioactive Compounds | Biological Mechanisms of Action Related to Neuroactivity |
|---|---|---|---|
| TANNINS |
(free-radical scavenging) (metal chelation) (pro-oxidative enzyme inhibition) (endogenous antioxidant system interaction) (inhibition of xanthine oxidase-induced Lipid peroxidation) [198][199]. |
ANTHOCYANIN | Delphinidin Cyanidin Anti-inflammatory, anti-Alzheimer, antitumor, and antioxidative properties by depleting the expression of cytokine markers [200]. |
Anti-inflammatory mechanisms, such as depletion of apoptotic cells [198]. |
|||
| QUERCETIN | MYRICETIN | Kaempferol Antitumor, anti-inflammation, and antioxidant properties are exercised via an antiproliferative mechanism in cells, attenuation mechanism against inflammation, and tumor growth factors [115][210]. |
|
|
|||
| ALKALOIDS | Vincristine Antineuroblastoma property exerted via the mechanisms truncating the glutathione metabolism [212]. Tetrandrine Anti-inflammatory and antitumor activities are linked to the calcium-channel blocking mechanism [213]. Skimmianine Anti-inflammatory property via the inhibition of nitric oxide production [214]. |
CATECHIN | |
| TERPENOIDS Paeoniflorin, Triptolidenol, Tripterine, Triptonide, Gindenoside, Oleanoic Acid |
Anti-inflammatory activity via interleukin-6 inhibition [217]. Anti-nociceptive, antioxidant and anti-inflammatory properties [218]. | RUTIN | |
| LIPID Omega-3 DHA |
Anti-inflammatory properties, Cell survival promotors [220]. |
Neuroprotectin D1 Lipid | Inhibition of apoptosis-related damage to DNA [220]. |
4. Concluding Remarks
Furthermore, the in silico computational model will help to potentially generate the biomechanistic and chemical information of natural products. In addition, the in silico and in vitro to in vivo frontiers offer more possibilities in bioactivity response simulation. Substantial in vitro, in vivo, and ex vivo evidence has been drawn about the capacity of naturally sourced bioactive principles (nutritive factors and secondary metabolites) to hijack or, even better, prevent the degenerative mechanisms that result in the widely known neurological diseases. Key neurologically relevant biochemical mechanisms, such as protein expression, up/downregulation, and gene targeting, provide insight into the modulatory fingerprints of these active molecules against neuropathogenesis.
NG108–15 glioblastoma
PGC-1/Nrf-2/HO-1 peroxisome proliferator-activated receptor coactivator/Nuclear factor2/Heme oxygenase
GFAP- glial fibrillary acidic protein
BDNF–TrkB-PI3K/Akt brain-derived neutrophic factor–tropomyosin-related kinase B/phosphatidylinositol
Bax/Bcl2 Bcl2 associated x/apoptosis regulator
TNF- α tumor necrosis factor-alpha
IL-1β interleukin-1β
COX-2 cyclooxygenase-2
P38MAPK p38 mitogen-activated protein kinase
PRP Prion Protein Peptide
HT22 hippocampus cell line
Aβ Amyloid Beta Peptide
TrKA tropomyosin receptor kinase A
ERK1/2 extracellular signal-related kinases 1/2
MPTP 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine
p-JNK p-Jun N-terminal kinase
p-ERK p-extracellular signal-regulated kinase protein
MBP maltose binding protein
DBH dopamine-β-hydroxylase
MAP2 microtubule-associated protein 2
GAP 43 growth-associated protein 43
PSD-95 postsynaptic density protein
KLK8 Kallikrein 8 gene
SH-SY5Y human neuroblastoma cell line
P53 tumor protein
β-secretase 1 Beta secretase 1
γ-secretase gamma secretase protein
NADH nicotinamide adenine dinucleotide hydrogen
NADPH nicotinamide adenine dinucleotide phosphate
PC12 pheochromocytoma cell line
GABA gamma-aminobutyric acid
ERβ estrogen receptor beta gene
NB69 Cellosaurus cell line
SNAP-25 synaptosomal-associated protein 25
CA1 Cortical Area 1
NF-kB nuclear factor-kappa B cells
STAT3 signal transducer and activator of transcription 3
ERK/MMP-9 extracellular signal-related kinases/matrix metallopeptidase-9
gp91phox 91-KD glycoprotein
TGF-β1 transforming growth factor
SK-N-SH neuroblastoma cell line
HIF-1α/LDHA hypoxia-inducible factor-1 alpha/Lactate dehydrogenase A gene
Nrf-2/Keap1 nuclear factor 2/Kelch-like ECH-associated protein 1
GSK-3β glycogen synthase kinase-3 beta
mTOR mammalian target of rapamycin
6-OHDA 6-hydroxydopamine
SOD-1 superoxide dismutase 1
TDP-43 TAR-DNA-binding protein 43
FUS/TLS fused in sarcoma/translocated in liposarcoma
anti-CHE anti-cholinesterase activity
ROS reactive oxygen species
HPLC High-Performance Liquid Chromatography
NMR Nuclear Magnetic Resonance
HNMR Proton Nuclear Magnetic Resonance
FTIR Fourier-Transform Infrared Spectroscopy
GAE gallic acid equivalent
QE quercetin equivalent
CE catechin equivalent
This entry is adapted from the peer-reviewed paper 10.3390/foods11152263
References
- Przedborski, S.; Vila, M.; Jackson-Lewis, V. Neurodegeneration: What is it and where are we? J. Clin. Investig. 2003, 111, 3–10.
- Johnston, M. Neurodegenerative disorders of childhood. In Nelson Textbook of Pediatrics, 20th ed.; Behrman, R.E., Kliegman, R.M., Jenson, H.B., Eds.; Elsevier: Philadelphia, PA, USA, 2016; p. 4264.
- Hardy, J. Pathways to Primary Neurodegenerative Disease. Mayo Clin. Proc. 1999, 74, 835–837.
- Chen, W.W.; Zhang, X.; Huang, W.J. Role of neuroinflammation in neurodegenerative diseases (Review). Mol. Med. Rep. 2016, 13, 3391–3396.
- Mobbs, C.V.; Hof, P.R. Body Composition and Aging; Interdisciplinary Top Gerontology; Karger: Basel, Switzerland, 2010; Volume 37, pp. 175–192.
- Lasonde, M.; Candel, S.; Hacker, J.; Quadrio-Curzio, A.; Onishi, T.; Ramakrishnan, V.; McNutt, M. The Challenge of Neurodegenerative Diseases in an Aging Population; G7 Academies’ Joint Statement; The Royal Society: London, UK, 2017; pp. 1–2.
- Taylor, J.P.; Hardy, J.; Fischbeck, K.H. Toxic proteins in neurodegenerative disease. Science 2002, 296, 1991–1995.
- Chen, H.; Chan, D.C. Mitochondrial dynamics- fusion, fission, movement, and mitophagy in neurodegenerative diseases. Hum. Mol. Genet. 2009, 18, R169–R176.
- Chin-Chan, M.; Navarro-Yepes, J.; Quintanilla-Vega, B. Environmental pollutants as risk factors for neurodegenerative disorders: Alzheimer and Parkinson diseases. Front. Cell. Neurosci. 2015, 124, 1–21.
- Dorsey, E.R.; Holloway, R.G.; Ravina, B.M. Biomarkers in Parkinson’s disease. Expert Rev. Neurother. 2006, 6, 823–831.
- Campbell, I.L.; Krucker, T.; Steffensen, S.; Akwa, Y.; Powell, H.C.; Lane, T.; Carr, D.J.; Gold, L.H.; Henriksen, S.J.; Siggins, G.R. Structural and functional neuropathology in transgenic mice with CNS expression of IFN-alpha. Brain Res. 1999, 835, 46–61.
- Ineichen, B. The epidemiology of dementia in Africa: A review. Soc. Sci. Med. 2000, 50, 1673–1677.
- Okubadejo, N.U.; Bower, J.H.; Rocca, W.A.; Maraganore, D.M. Parkinson’s disease in Africa. A systematic review of epidemiologic and genetic studies. Mov. Disord. 2006, 21, 2150–2156.
- Cilia, R.; Akpalu, A.; Cham, M.; Bonetti, A.; Amboni, M.; Faceli, E.; Pezzoli, G. Parkinson’s disease in sub-Saharan Africa: Step-by-step into the challenge. Neurodegener. Dis. Manag. 2011, 1, 193–202.
- George-Carey, R.; Adeloye, D.; Chan, K.Y.; Paul, A.; Kolcić, I.; Campbell, H.; Rudan, I. An estimate of the prevalence of dementia in Africa: A systematic analysis. J. Glob. Health 2012, 2, 1–13.
- Marin, B.; Kacem, I.; Diagana, M.; Boulesteix, M.; Gouider, R.; Preux, P.M.; Couratier, P. Juvenile, and adult-onset ALS/MND among Africans: Incidence, phenotype, survival: A review. ALS 2012, 13, 276–283.
- Onyike, C.U.; Diehl-Schmid, J. The epidemiology of frontotemporal dementia. Int. Rev. Psychiatry 2013, 25, 130–137.
- Bower, J.H.; Maraganore, D.M.; McDonnell, S.K.; Rocca, W.A. Incidence of progressive supranuclear palsy and multiple system atrophy in Olmsted County, Minnesota, 1976 to 1990. Neurology 1997, 49, 1284–1288.
- Coon, E.A.; Sletten, D.M.; Suarez, M.D.; Mandrekar, J.N.; Ahlskog, J.E.; Bower, J.H.; Matsumoto, J.Y.; Silber, M.H.; Benarroch, E.E.; Fealey, R.D.; et al. Clinical features and autonomic testing predict survival in multiple system atrophy. Brain 2015, 138, 3623–3631.
- Köllensperger, M.; Geser, F.; Ndayisaba, J.; Boesch, S.; Seppi, K.; Ostergaard, K.; Dupont, E.; Cardozo, A.; Tolosa, E.; Abele, M.; et al. Presentation, diagnosis, and management of multiple system atrophy in Europe: Final analysis of the European multiple system atrophy registry. Mov. Disord. 2010, 25, 2604–2612.
- Tan, S.J.; Ismail, I.S. Potency of Selected Berries, Grapes, and Citrus Fruit as Neuroprotective Agents. Evid. Based Complement. Alternat. Med. 2020, 2020, 1–12.
- Xia, E.Q.; Deng, G.F.; Guo, Y.J.; Li, H.B. Biological activities of polyphenols from grapes. Int. J. Mol. Sci. 2010, 11, 622–646.
- Fu, L.; Xu, B.T.; Xu, X.R.; Gan, R.Y.; Zhang, Y.; Xia, E.Q.; Li, H.B. Antioxidant capacities and total phenolic contents of 62 fruits. Food Chem. 2011, 129, 345–350.
- Deng, G.F.; Shen, C.; Xu, X.R.; Kuang, R.D.; Guo, Y.J.; Zeng, L.S.; Gao, L.L.; Lin, X.; Xie, J.F.; Xia, E.Q.; et al. Potential of Fruit Wastes as Natural Resources of Bioactive Compounds. Int. J. Mol. Sci. 2012, 13, 8308–8323.
- Li, F.; Li, S.; Li, H.B.; Deng, G.F.; Ling, W.H.; Wu, S.; Xu, X.R.; Chen, F. Antiproliferative activity of peels, pulps, and seeds of 61 fruits. J. Funct. Foods 2013, 5, 1298–1309.
- Ishurda, O.; John, F.K. The anti-cancer activity of polysaccharides prepared from Libyan dates (Phoenix dactylifera L.). Carbohydr. Polym. 2005, 59, 531–535.
- Al-Qarawi, A.A.; Abdel-Rahman, H.; Mousa, H.M.; Ali, B.H.; El-Mougy, S.A. Nephroprotective action of Phoenix dactylifera in Gentamicin-Induced Nephrotoxicity. Pharm. Biol. 2008, 46, 227–230.
- Saafi, E.B.; Louedi, M.; Elfeki, A.; Zakhama, A.; Najjar, M.F.; Hammami, M.; Achour, L. Protective effect of date palm fruit extract (Phoenix dactylifera L.) on dimethoate induced oxidative stress in rat liver. Exp. Toxicol. Pathol. 2011, 63, 433–441.
- Kasi, P.B.; Kotormán, M. Avocado juice prevents the formation of Trypsin amyloid-like fibrils in aqueous ethanol. Nat. Prod. Commun. 2019, 14, 1–5.
- Joseph, J.A.; Shukitt-Hale, B.; Denisova, N.A.; Prior, R.L.; Cao, G.; Martin, A.; Taglialatela, G.; Bickford, P.C. Long-term dietary strawberry, spinach, or vitamin E supplementation retards the onset of age-related neuronal signal-transduction and cognitive behavioral deficits. J. Neurosci. 1998, 18, 8047–8055.
- Joseph, J.A.; Shukitt-Hale, B.; Denisova, N.A.; Bielinski, D.; Martin, A.; McEwen, J.J.; Bickford, J.J. Reversals of age-related declines in neuronal signal transduction, cognitive, and motor behavioral deficits with blueberry, spinach, or strawberry dietary supplementation. J. Neurosci. 1999, 19, 8114–8121.
- Gambino, G.; Allegra, M.; Sardo, P.; Attanzio, A.; Tesoriere, L.; Livrea, M.A.; Ferraro, G.; Carletti, F. Brain distribution and modulation of neuronal excitability by indicaxanthin from Opuntia ficus indica administered at nutritionally relevant amounts. Front. Aging Neurosci. 2018, 10, 1–11.
- Oduje, A.A.; Oboh, G.; Ayodele, A.J.; Stephen, A.A. Assessment of the nutritional, antinutritional, and antioxidant capacity of unripe, ripe, and overripe plantain (Musa paradisiaca) peel. Int. J. Curr. Adv. Res. 2015, 3, 63–72.
- Ogbonna, O.A.; Izundu, A.I.; Okoye, N.H.; Ikeyi, A.P. Phytochemical compositions of fruits of three Musa species at three stages of development. IOSR J. Pharm. 2016, 11, 48–59.
- Aboul-Enein, A.M.; Salama, Z.A.; Gaafar, A.A.; Aly, H.F.; Abou-Elella, F.; Ahmed, H.A. Identification of phenolic compounds from the banana peel (Musa paradisiaca L.) as antioxidant and antimicrobial agents. J. Chem. Pharm. 2016, 8, 46–55.
- Agama-Acevedo, E.; Sañudo-Barajas, J.A.; Vélez De La Rocha, R.; González-Aguilar, G.A.; Bello-Peréz, L.A. Potential of plantain peels flour (Musa paradisiaca L.) as a source of dietary fiber and antioxidant compound. CYTA J. Food 2016, 14, 117–123.
- Aina, O.O.; Oyedeji, M.B.; Adegboyega, D.A.; Owoloja, A.O. Phytochemical screening of some selected banana peels of Musa acuminata species. Int. J. Agric. 2019, 4, 68–78.
- Boua, B.B.; Ouattara, D.; Traoré, L.; Mamyrbekova-Békro, J.A.; Békro, Y.A. Effect of domestic cooking on the total phenolic, flavonoid and condensed tannin content from plantain of Côte d’Ivoire. J. Mater. Environ. Sci. 2020, 11, 396–403.
- Ashafaq, M.; Tabassum, H.; Vishnoi, S.; Salman, M.; Raisuddin, S.; Parvez, S. Tannic acid alleviates lead acetate-induced neurochemical perturbations in rat brain. Neurosci. Lett. 2016, 617, 94–100.
- Salman, M.; Tabassum, H.; Parvez, S. Nrf2/HO-1 mediates the neuroprotective effects of pramipexole by attenuating oxidative damage and mitochondrial perturbation after traumatic brain injury in rats. Dis. Model Mech. 2020, 13, 1–11.
- Sulaiman, S.F.; Yusoff, N.A.M.; Eldeen, I.M.; Seow, E.M.; Sajak, A.A.; Supriatno; Ooi, K.L. Correlation between total phenolic and mineral contents with antioxidant activity of eight Malaysian bananas (Musa sp.). J. Food Compost. Anal. 2011, 24, 1–10.
- Chauhan, A.; Nagar, A.; Bala, K.; Sharma, Y. Comparative study of different parts of fruits of Musa Sp. on the basis of their antioxidant activity. Der Pharm. Lett. 2016, 8, 88–100.
- Al Amri, F.S.; Hossain, M.A. Comparison of total phenols, flavonoids, and antioxidant potential of local and imported ripe bananas. Egypt. J. Basic Appl. Sci. 2018, 5, 245–251.
- Kamaayi, F.; Baah, F.A.; Ansah, F.O. Phenolic content, polyphenol oxidase activity, and antioxidant scavenging activity in three species of plantain in Ghana. Int. J. Sci. Res. 2020, 10, 551–557.
- Fatemeh, S.R.; Saifullah, R.; Abbas, F.M.A.; Azhar, M.E. Total phenolics, flavonoids, and antioxidant activity of banana pulp and peel flours: Influence of variety and stage of ripeness. Int. Food Res. J. 2012, 19, 1041–1046.
- Oyeyinka, B.O.; Afolayan, A.J. Comparative and Correlational evaluation of the phytochemical constituents and antioxidant activity of Musa sinensis L. and Musa paradisiaca L. fruit compartments (Musaceae). Sci. World J. 2020, 2020, 1–12.
- Ssonko, U.L.; Muranga, F.I. Phenolic compounds identification and antioxidant activity in bananas of AAB and ABB genomes grown in Uganda. J. Health Popul. Nutr. 2020, 4, 1–8.
- Kevers, C.; Falkowski, M.; Tabart, J.; Defraigne, J.O.; Dommes, J.; Pincemail, J. Evolution of antioxidant capacity during storage of selected fruits and vegetables. J. Agric. Food Chem. 2007, 55, 8596–8603.
- Yao, J.; Irwin, R.; Chen, S.; Hamilton, R.; Cadenas, E.; Brinton, R.D. Ovarian hormone loss induces bioenergetic deficits and mitochondrial β-amyloid. Neurobiol. Aging 2012, 33, 1507–1521.
- Khan, I.; Bahuguna, A.; Bhardwaj, M.; Khaket, T.P.; Kang, S.C. Carvacrol nano-emulsion evokes cell cycle arrest, apoptosis induction, and autophagy inhibition in a doxorubicin-resistant A549 cell line. Artif. Cells Nanomed. Biotechnol. 2018, 46, S664–S675.
- Tsamo, C.V.P.; Herent, M.F.; Tomekpe, K.; Emaga, T.H.; Quetin-Leclercq, J.; Rogez, H.; Larondelle, Y.; Andre, C. Phenolic profiling in the pulp and peel of nine plantain cultivars (Musa sp.). Food Chem. 2015, 167, 197–204.
- Jiménez-Aliaga, K.; Bermejo-Bescós, P.; Benedí, J.; Martín-Aragón, S. Quercetin and rutin exhibit antiamyloidogenic and fibril-disaggregating effects in vitro and potent antioxidant activity in APPswe cells. Life Sci. 2011, 89, 939–945.
- Wang, Y.; Szretter, K.J.; Vermi, W.; Gilfillan, S.; Rossini, C.; Cella, M.; Barrow, A.D.; Diamond, M.S.; Colonna, M. IL-34 is a tissue-restricted ligand of CSF1R required for the development of Langerhans cells and microglia. Nat. Immunol. 2012, 13, 753–760.
- Javed, H.; Khan, M.M.; Ahmad, A.; Vaibhav, K.; Ahmad, M.E.; Khan, A.; Ashafaq, M.; Islam, F.; Siddiqui, M.S.; Safhi, M.M.; et al. Rutin prevents cognitive impairments by ameliorating oxidative stress and neuroinflammation in the rat model of sporadic dementia of Alzheimer’s type. Neuroscience 2012, 210, 340–352.
- Na, J.Y.; Kim, S.; Song, K.; Kwon, J. Rutin alleviates Prion peptide-induced cell death through inhibiting apoptotic pathway activation in dopaminergic neuronal cells. Cell. Mol. Neurobiol. 2014, 34, 1071–1079.
- Xu, Y.; Zhang, M.; Ramos, C.A.; Durett, A.; Liu, E.; Dakhova, O.; Liu, H.; Creighton, C.J.; Gee, A.P.; Heslop, H.E.; et al. Closely related T-memory stem cells correlate with in vivo expansion of CAR.CD19-T cells and are preserved by IL-7 and IL-15. Blood 2014, 123, 3750–3759.
- Ganeshpurkar, A.; Saluja, A.K. The pharmacological potential of Rutin. Saudi Pharm. J. 2017, 25, 149–164.
- Song, H.; Zhang, X.; Wang, W.; Liu, R.; Zhao, K.; Liu, M.; Gong, W.; Ning, B. Neuroprotective mechanisms of rutin for spinal cord injury through anti-oxidation and anti-inflammation and inhibition of the p38 mitogen-activated protein kinase pathway. Neural Regen. Res. 2018, 13, 128–134.
- Adepoju, O.T.; Sunday, B.E.; Folaranmi, O.A. Nutrient composition and contribution of plantain (Musa paradisiaca) products to dietary diversity of Nigerian consumers. Afr. J. Biotechnol. 2012, 11, 13601–13605.
- Okareh, O.T.; Adeolu, A.T.; Adepoju, O.T. Proximate and mineral composition of plantain (Musa paradisiaca) wastes flour; a potential nutrients source in the formulation of animal feeds. Afr. J. Food Sci. 2015, 6, 53–57.
- Hapsari, L.; Lestari, D.A. Fruit characteristics and nutrient values of four Indonesian banana cultivars (Musa spp.) at different genomic groups. AGRAVITA J. Agric. Sci. 2016, 38, 303–311.
- Siji, S.; Nandini, P.V. Chemical and nutrient composition of selected banana varieties of Kerala. Int. J. Adv. Eng. Manag. Sci. 2017, 3, 401–404.
- Ho, Y.S.; Yu, M.S.; Yang, X.F.; So, K.F.; Yuen, W.H.; Chang, R.C.C. Neuroprotective effects of polysaccharides from wolfberry, the fruits of Lycium barbarum, against homocysteine- induced toxicity in rat cortical neurons. J. Alzheimer’s Dis. 2010, 19, 813–827.
- Chiricozzi, A.; De Simone, C.; Fossati, B.; Peris, K. Emerging treatment options for the treatment of moderate to severe plaque psoriasis and psoriatic arthritis: Evaluating bimekizumab and its therapeutic potential. Psoriasis Targets Ther. 2019, 9, 29–35.
- Hasanah, R.; Daningsih, E.; Titin, T. The analysis of nutrient and fiber content of banana (Musa paradisiaca) sold in Pontianak, Indonesia. Biofarmasi J. Nat. Prod. Biochem. 2017, 15, 21–25.
- Abubakar, U.S.; Yusuf, K.M.; Safiyanu, I.; Abdullahi, S.; Saidu, S.R.; Abdu, G.T.; Indee, A.M. Proximate and mineral composition of corn cob, banana, and plantain peels. Int. J. Food Sci. 2016, 1, 25–27.
- Adamu, A.S.; Ojo, I.O.; Oyetunde, J.G. Evaluation of nutritional values in ripe, unripe, boiled, and roasted plantain (Musa paradisiaca L.) pulp and peel. Eur. J. Basic Appl. Sci. 2017, 4, 9–12.
- Shadrach, I.; Banji, A.; Adebayo, O. Nutraceutical potential of ripe and unripe plantain peels: A comparative study. Chem. Int. 2020, 6, 83–90.
- Kirkland, A.E.; Sarlo, G.L.; Holton, K.F. The role of magnesium in neurological disorders. Nutrients 2018, 10, 730.
- Annor, G.A.; Asamoah-Bonti, P.; Sakyi-Dawson, E. Fruit physical characteristics, proximate, mineral and starch characterization of FHIA 19 and FHIA 20 plantain and FHIA 03 cooking banana hybrids. SpringerPlus 2016, 5, 796.
- Seth, R.; Corniola, R.S.; Gower-Winter, S.D.; Morgan, T.J., Jr.; Bishop, B.; Levenson, C.W. Zinc deficiency induces apoptosis via mitochondrial p53- and caspase-dependent pathways in human neuronal precursor cells. J. Trace Elem. Med. Biol. 2015, 30, 59–65.
- Chen, N.N.; Zhao, D.J.; Sun, Y.X.; Wang, D.D.; Ni, H. Long-Term Effects of Zinc Deficiency and Zinc Supplementation on Developmental Seizure-Induced Brain Damage and the Underlying GPR39/ZnT-3 and MBP Expression in the Hippocampus. Front. Neurosci. 2019, 13, 1–10.
- Marasco, D.; Vicidomini, C.; Krupa, P.; Cioffi, F.; Huy, P.D.Q.; Li, M.S.; Florio, D.; Broersen, K.; Francesca De Pandis, M.; Roviello, G.N. Plant isoquinoline alkaloids as potential neuro drugs: A comparative study of the effects of benzophenanthridine and berberine-based compounds on β-amyloid aggregation. Chem. Biol. Interact. 2021, 334, 109300.
- Konar, A.; Shah, N.; Singh, R.; Saxena, N.; Kaul, S.C.; Wadhwa, R.; Thakur, M.K. Protective Role of Ashwagandha leaf extract and its component Withanone on scopolamine-induced changes in the brain and brain-derived cells. PLoS ONE 2011, 6, e27265.
- Konar, A.; Gupta, R.; Shukla, R.K.; Maloney, B.; Khanna, V.K.; Wadhwa, R.; Lahiri, D.K.; Thakur, M.K. M1 muscarinic receptor is a key target of neuroprotection, neuroregeneration, and memory recovery by i-Extract from Withania somnifera. Sci. Rep. 2019, 9, 13990.
- Oyeyinka, B.O.; Afolayan, A.J. Comparative Evaluation of the Nutritive, Mineral, and Antinutritive Composition of Musa sinensis L. (Banana) and Musa paradisiaca L. (Plantain). Plants 2019, 8, 598.
- Oyeyemi, S.D.; Otoide, J.E.; Obembe, M.O.; Ogunleye, B.J. Nutritional and phytochemical assessment of Musa sapientum var. velutina. The Pharm. Chem. J. 2019, 6, 56–64.
- Chen, Y.; Sun, H.; Huang, L.; Li, J.; Zhou, W.; Chang, J. Neuroprotective effect of Radix trichosanthis saponins on subarachnoid hemorrhage. Evid. Based Complement. Alternat. Med. 2015, 2015, 1–10.
- Khawas, P.; Das, A.J.; Sit, N.; Badwaik, L.S.; Deka, S.C. Nutritional composition of culinary Musa ABB at Different Stages of Development. Am. J. Food Technol. 2014, 2, 80–87.
- Anekonda, T.S.; Quinn, J.F.; Harris, C.; Frahler, K.; Wadsworth, T.L.; Woltjer, R.L. L-type voltage-gated calcium channel blockade with isradipine as a therapeutic strategy for Alzheimer’s disease. Neurobiol. Dis. 2011, 41, 61–70.
- Thinh, B.B.; Trong, L.V.; Lam, L.T.; Hien, V.T.T. Nutritional value of persimmon, banana, lemon, and longan cultivated in Northern Vietnam. IOP Conf. Ser. Earth Environ. Sci. 2021, 640, 022030.
- Velumani, S. Phytochemical screening and antioxidant activity of banana peel. Int. J. Adv. Res. Innov. Ideas Educ. 2016, 2, 91–102.
- Kim, S.M.; Chung, M.J.; Ha, T.J.; Choi, H.N.; Jang, S.J.; Kim, S.O.; Chun, M.H.; Do, S.I.; Choo, Y.K.; Park, Y.I. Neuroprotective effects of black soybean anthocyanins via inactivation of ASK1-JNK/p38 pathways and mobilization of cellular sialic acids. Life Sci. 2012, 90, 874–882.
- Lau, F.C.; Joseph, J.A.; McDonald, J.E.; Kalt, W. Attenuation of iNOS and COX2 by blueberry polyphenols is mediated through the suppression of NF-κB activation. J. Funct. Foods 2009, 1, 274–283.
- Ma, C.; Zhao, L.L.; Zhao, H.J.; Cui, J.W.; Li, W.; Wang, N.Y. Lentivirus-mediated MDA7/IL24 expression inhibits the proliferation of hepatocellular carcinoma cells. Mol. Med. Rep. 2018, 17, 5764–5773.
- Agoreyo, B.O.; Agoreyo, F.O.; Omigie, M.I. Antioxidant activity, phytochemical and antioxidant levels of Musa paradisciaca L. and Musa sapientum L. at various ripening stages. Eur. Food Res. Technol. 2017, 5, 41–59.
- Nakaso, K.; Tajima, N.; Horikoshi, Y.; Nakasone, M.; Hanaki, T.; Kamizaki, K.; Matsura, T. The estrogen receptor β-PI3K/Akt pathway mediates the cytoprotective effects of tocotrienol in a cellular Parkinson’s disease model. Biochim. Biophys. Acta BBA Mol. Basis Dis. 2014, 1842, 1303–1312.
- Al-Ahmary, K.M. Selenium content in selected foods from the Saudi Arabia market and estimation of the daily intake. Arab. J. Chem. 2009, 2, 95–99.
- Choi, Y.; Kim, J.; Lee, H.S.; Kim, C.; Hwang, I.K.; Park, H.K.; Oh, C.H. Selenium content in representative Korean foods. J. Food Compos. Anal. 2009, 22, 117–122.
- Al-Othman, A.M.; Al-Othman, Z.A.; El-Desoky, G.E.; Aboul-Soud, M.A.M.; Habila, M.A.; Giesy, J.P. Daily intake of selenium and concentrations in blood of residents of Riyadh City, Saudi Arabia. Environ. Geochem. Health 2012, 34, 417–431.
- Moatkhef, F.; Ismail, H.; Agamy, N.; Aborhyem, S. Quantitative determination of selenium in the most common food items sold in Egypt. J. Egypt. Public Health Assoc. 2020, 95, 1–9.
- Naziroğlu, M.; Kutluhan, S.; Yilmaz, M. Selenium and topiramate modulate brain microsomal oxidative stress values, Ca2+-ATPase activity, and EEG records in pentylenetetrazol-induced seizures in rats. J. Membr. Biol. 2008, 225, 39–49.
- Naziroğlu, M.; Şenol, N.; Ghazizadeh, V.; Yyürüker, V. Neuroprotection induced by N-acetylcysteine and selenium against traumatic brain injury-induced apoptosis and calcium entry in the hippocampus of the rat. Cell. Mol. Neurobiol. 2014, 34, 895–903.
- Klingberg, S.; Andersson, H.; Mulligan, A.; Bhaniani, A.; Welch, A.; Bingham, S.; Khaw, K.T.; Andersson, S.; Ellegard, L. Food sources of plant sterols in the EPIC Norfolk population. Eur. J. Clin. Nutr. 2008, 62, 695–703.
- Rui, X.; Wenfang, L.; Jing, C.; Meng, C.; Chengcheng, D.; Jiqu, X.; Shuang, R. Neuroprotective effects of phytosterol esters against high cholesterol-induced cognitive deficits in the aged rat. Food Funct. 2017, 8, 1323–1332.
- Kopylov, A.T.; Malsagova, K.A.; Stepanov, A.A.; Kaysheva, A.L. Diversity of Plant Sterols Metabolism: The impact on human health, sport, and accumulation of contaminating sterols. Nutrients 2021, 13, 1623.
- Uzairu, S.M.; Kano, M.A. Assessment of phytochemical and mineral composition of unripe and ripe plantain (Musa paradisiaca) peels. Afr. J. Food Sci. 2021, 15, 107–112.
- Kasture, V.S.; Deshmukh, V.K.; Chopde, C.T. Anticonvulsant and behavioral actions of triterpene isolated from Rubia cordifolia Linn. Indian J. Exp. Biol. 2000, 38, 675–680.
- Yu, L.; Chen, C.; Wang, L.F.; Kuang, X.; Liu, K.; Zhang, H.; Du, J.R. Neuroprotective effect of Kaempferol glycosides against brain injury and neuroinflammation by inhibiting the activation of NF-κB and STAT3 in transient focal stroke. PLoS ONE 2013, 8, e55839.
- Yang, J.; Zeng, Z.; Wu, T.; Yang, Z.; Liu, B.; Lan, T. Emodin attenuates high glucose-induced TGF-β1 and fibronectin expression in mesangial cells through inhibition of the NF-κB pathway. Exp. Cell Res. 2013, 319, 3182–3189.
- Wang, T.; Fan, X.; Tang, T.; Fan, R.; Zhang, C.; Huang, Z.; Peng, W.; Gan, P.; Xiong, X.; Huang, W.; et al. Rhein, and rhubarb similarly protect the blood-brain barrier after experimental traumatic brain injury via gp91phox subunit of NADPH oxidase/ROS/ERK/ MMP-9 signaling pathway. Sci. Rep. 2016, 6, 37098.
- Aurore, G.; Parfait, B.; Fahrasmane, L. Bananas, raw materials for making processed food products. Trends Food Sci. Technol. 2009, 20, 78–91.
- Chen, Y.M.; Li, H.; Chiu, Y.S.; Huang, C.C.; Chen, W.C. Supplementation of L-arginine, L- glutamine, Vitamin C, Vitamin E, folic acid, and green tea extract enhances serum nitric oxide content and anti-fatigue activity in mice. Evid. Based Complement. Alternat. Med. 2020, 2020, 1–10.
- Prakash, A.; Kumar, A. Implicating the role of lycopene in the restoration of mitochondrial enzymes and BDNF levels in β-amyloid induced Alzheimer’s disease. Eur. J. Pharmacol. 2014, 741, 104–111.
- Hong, Y.G.; Roh, S.; Paik, D.; Jeong, S. Development of a reporter system for in vivo monitoring of γ-secretase activity in Drosophila. Mol. Cells 2017, 40, 73–81.
- Heng, Z.; Sheng, O.; Yan, S.; Lu, H.; Motorykin, I.; Gao, H.; Li, C.; Yang, Q.; Hu, C.; Kuang, R.; et al. Carotenoid profiling in the peel and pulp of 36 selected Musa varieties. Food Sci. Technol. Res. 2017, 23, 603–611.
- Villaverde, J.J.; Oliveira, L.; Vilela, C.; Domingues, R.M.; Freitas, N.; Cordeiro, N.; Freire, C.S.R.; Silvestre, A.J.D. High valuable compounds from the unripe peel of several Musa species cultivated in Madeira Island (Portugal). Ind. Crops Prod. 2013, 42, 507–512.
- Shi, C.; Liu, J.; Wu, F.; Zhu, X.; Yew, D.T.; Xu, J. β-sitosterol inhibits high cholesterol-induced platelet β-amyloid release. J. Bioenerg. Biomembr. 2011, 43, 691–697.
- Shi, C.; Wu, F.; Zhu, X.C.; Xu, J. Incorporation of beta-sitosterol into the membrane increases resistance to oxidative stress and lipid peroxidation via estrogen receptor-mediated PI3K/GSK3beta signaling. Biochim. Biophys. Acta 2013, 1830, 2538–2544.
- Mordi, R.C.; Fadiaro, A.E.; Owoeye, T.F.; Olanrewaju, I.O.; Uzoamaka, G.C.; Olorunshola, S.J. Identification by GC-MS of the components of oils of banana peels extract, phytochemical and antimicrobial analyses. Res. J. Phytochem. 2016, 10, 39–44.
- Hsieh, P.F.; Hou, C.W.; Yao, P.W.; Wu, S.P.; Peng, Y.F.; Shen, M.L.; Lin, C.H.; Chao, Y.Y.; Chang, M.H.; Jeng, K.C. Sesamin ameliorates oxidative stress and mortality in kainic acid-induced status epilepticus by inhibition of MAPK and COX-2 activation. J. Neuroinflamm. 2011, 8, 57.
- Xu, Z.; Liu, Y.; Yang, D.; Yuan, F.; Ding, J.; Chen, H.; Tian, H. Sesamin protects SH-SY5Y cells against mechanical stretch injury and promoting cell survival. BMC Neurosci. 2017, 18, 57.
- Udomruk, S.; Kaewmool, C.; Phitak, T.; Pothacharoen, P.; Kongtawelert, P. Sesamin promotes neurite outgrowth under insufficient nerve growth factor condition in PC12 cells through the ERK1/2 pathway and SIRT1 modulation. Evid. Based Complement. Alternat. Med. 2020, 2020, 1–12.
- Singh, B.; Singh, J.P.; Kaur, A.; Singh, N. Bioactive compounds in banana and their associated health benefits—A review. Food Chem. 2016, 206, 1–11.
- Zhang, N.; Feng, H.; Liao, H.H.; Chen, S.; Yang, Z.; Deng, W.; Tang, Q.Z. Myricetin attenuated LPS induced cardiac injury in vivo and in vitro. Phytother. Res. 2018, 32, 459–470.
- Rebello, L.P.G.; Ramos, A.M.; Pertuzatti, P.B.; Barcia, M.T.; Muñoz, N.C.; Hermosín-Gutiérrez, I. Flour of banana (Musa AAA) peel as a source of antioxidant phenolic compounds. Food Res. Int. 2014, 55, 397–403.
- Rinaldo, D. Carbohydrate and bioactive compounds composition of starchy tropical fruits and tubers, in relation to pre and postharvest conditions: A review. J. Food Sci. 2020, 85, 249–259.
- Vlachos, G.S.; Scarmeas, N. Dietary interventions in mild cognitive impairment and dementia. Dialogues Clin. Neurosci. 2019, 21, 69–82.
- Singh, P.; Sivanandama, T.M.; Konar, A.; Thakur, M.K. Role of nutraceuticals in cognition during aging and related disorders. Neurochem. Int. 2021, 143, 104928.
- Tan, H.P.; Wong, D.Z.H.; Ling, S.K.; Chuah, C.H.; AbdulKadir, H. Neuroprotective activity of galloylated cyanogenic glucosides and hydrolyzable tannins isolated from leaves of Phyllagathis rotundifolia. Fitoterapia 2012, 83, 223–229.
- Winiarska-Mieczan, A. Protective effect of tannic acid on the brain of adult rats exposed to cadmium and lead. Environ. Toxicol. Pharmacol. 2013, 36, 9–18.
- Calou, I.; Bandeira, M.A.; Aguiar-Galvão, W.; Cerqueira, G.; Siqueira, R.; Neves, K.R.; Brito, G.A.; Viana, G. Neuroprotective properties of a standardized extract from Myracrodruon urundeuva Fr. All. (Aroeira-Do-Sertão), as evaluated by a Parkinson’s disease model in rats. Parkinsons Dis. 2014, 2014, 1–12.
- Zengin, G.; Locatelli, M.; Carradori, S.; Mocan, A.M.; Aktumsek, A. Total phenolics, flavonoids, condensed tannins content of eight Centaurea species and their broad inhibitory activities against cholinesterase, tyrosinase, α-amylase, and α-glucosidase. Not. Bot. Horti Agrobot. 2016, 44, 195–200.
- Liu, X.; Yan, Y.; Li, F.; Zhang, D. Fruit and vegetable consumption and the risk of depression: A meta-analysis. Nutrition 2016, 32, 296–302.
- Türedi, S.; Yuluğ, E.; Alver, A.; Bodur, A.; İnce, I. A morphological and biochemical evaluation of the effects of quercetin on experimental sciatic nerve damage in rats. Exp. Ther. Med. 2018, 15, 3215–3224.
- Sharma, D.R.; Wani, W.Y.; Sunkaria, A.; Kandimalla, R.J.L.; Verma, D.; Cameotra, S.S.; Gill, K.D. Quercetin protects against chronic aluminum-induced oxidative stress and ensuing biochemical, cholinergic, and neurobehavioral impairments in rats. Neurotox. Res. 2013, 23, 336–357.
- Wassem, M.; Parvez, S. Neuroprotective activities of curcumin and quercetin with potential relevance to mitochondrial dysfunction induced by oxaliplatin. Protoplasma 2015, 253, 417–430.
- La Casa, C.; Villegas, I.; Alarcón de la Lastra, C.; Motilva, V.; Martín Calero, M.J. Evidence for protective and antioxidant properties of rutin, a natural flavone, against ethanol-induced gastric lesions. J. Ethnopharmacol. 2000, 71, 45–53.
- Schwedhelm, E.; Maas, R.; Troost, R.; Boger, R.H. Clinical pharmacokinetics of antioxidants and their impact on systemic oxidative stress. Clin. Pharmacokinet. 2003, 42, 437–459.
- Nassiri-Asl, M.; Mortazavi, S.R.; Samiee-Rad, F.; Zangivand, A.A.; Safdari, F.; Saroukhani, S.; Abbasi, E. The effects of rutin on the development of pentylenetetrazole kindling and memory retrieval in rats. Epilepsy Behav. 2010, 18, 50–53.
- Richetti, S.K.; Blank, M.; Capiotti, K.M.; Piato, A.L.; Bogo, M.R.; Vianna, M.R.; Bonan, C.D. Quercetin and rutin prevent scopolamine-induced memory impairment in zebrafish. Behav. Brain Res. 2011, 217, 10–15.
- Ramalingayya, G.V.; Cheruku, S.P.; Nayak, P.G.; Kishore, A.; Shenoy, R.; Rao, C.M.; Krishnadas, N. Rutin protects against neuronal damage in vitro and ameliorates doxorubicin-induced memory deficits in vivo in Wistar rats. Drug Des. Devel. Ther. 2017, 11, 1011–1026.
- Kumar, A.; Rinwa, P.; Dhar, H. Possible nitric oxide modulation in the protective effects of rutin against experimental head trauma-induced cognitive deficits: Behavioral, biochemical, and molecular correlates. J. Surg. Res. 2014, 188, 268–279.
- Robinson, D.T.; Martin, C.R. Fatty acid requirements for the preterm infant. Semin. Fetal Neonatal Med. 2017, 22, 8–14.
- Lo Van, A.; Sakayori, N.; Hachem, M.; Belkouch, M.; Picq, M.; Fourmaux, B.; Lagarde, M.; Osumi, N.; Bernoud-Hubac, N. Targeting the brain with a neuroprotective Omega-3 fatty acid to enhance neurogenesis in the hypoxic condition in culture. Mol. Neurobiol. 2018, 56, 986–999.
- Calderon, F.; Kim, H.Y. Docosahexaenoic acid promotes neurite growth in hippocampal neurons. J. Neurochem. 2004, 90, 979–988.
- Barker-Haliski, M.L.; Dahle, E.J.; Heck, T.D.; Pruess, T.H.; Vanegas, F.; Wilcox, K.S.; White, H.S. Evaluating an etiologically relevant platform for therapy development for temporal lobe epilepsy: Effects of carbamazepine and valproic acid on acute seizures and chronic behavioral comorbidities in the Theiler’s murine encephalomyelitis virus mouse model. J. Pharmacol. Exp. Ther. 2015, 353, 318–329.
- Gröber, U.; Schmidt, J.; Kisters, K. Magnesium in Prevention and Therapy. Nutrients 2015, 7, 8199–8226.
- Clerc, P.; Young, C.A.; Bordt, E.A.; Grigore, A.M.; Fiskum, G.; Polster, B.M. Magnesium sulfate protects against the bioenergetic consequences of chronic glutamate receptor stimulation. PLoS ONE 2013, 8, e79982.
- Lambuk, L.; Jafri, A.J.A.; Arfuzir, N.N.N.; Iezhitsa, I.; Agarwal, R.; Rozali, K.N.B.; Agarwal, P.; Bakar, N.S.; Kutty, M.K.; Yusof, A.P.; et al. Neuroprotective effect of Magnesium acetyltaurate against NMDA-induced excitotoxicity in rat retina. Neurotox. Res. 2017, 31, 31–45.
- Adamo, A.M.; Zago, M.P.; Mackenzie, G.G.; Aimo, L.; Keen, C.L.; Keenan, A.; Oteiza, P.I. The role of zinc in the modulation of neuronal proliferation and apoptosis. Neurotox. Res. 2010, 17, 1–14.
- Osredkar, J.; Sustar, N. Copper and Zinc, Biological Role and Significance of Copper/Zinc Imbalance. J. Clin. Toxicol. S 2011, 3, 1–18.
- Kaler, S.G.; Holmes, C.S.; Goldstein, D.S.; Tang, J.; Godwin, S.C.; Donsante, A.; Liew, C.J.; Sato, S.; Patronas, N. Neonatal diagnosis, and treatment of Menkes disease. N. Engl. J. Med. 2008, 358, 605–614.
- Kaler, S.G. ATP7A-related copper transport diseases-emerging concepts and future trends. Nat. Rev. Neurol. 2011, 7, 15–29.
- Rubio-Osornio, M.; Montes, S.; Perez-Severiano, F.; Aguilera, P.; Floriano-Sanchez, E.; Monroy-Noyola, A.; Rubio, C.; Rios, C. Copper reduces striatal protein nitration and tyrosine hydroxylase inactivation induced by MPP+ in rats. Neurochem. Int. 2009, 54, 447–451.
- Sehgal, N.; Gupta, A.; Valli, R.K.; Joshi, S.D.; Mills, J.T.; Hamel, E.; Khanna, P.; Jain, S.C.; Thakur, S.S.; Ravindranath, V. Withania somnifera reverses Alzheimer’s disease pathology by enhancing low-density lipoprotein receptor-related protein in the liver. Proc. Natl. Acad. Sci. USA 2012, 109, 3510–3515.
- Liu, Y.; Liu, X.; Hua, W.; Wei, Q.; Fang, X.; Zhao, Z.; Chun, G.; Chao, L.; Chen, C.; Yifu, T.; et al. Berberine inhibits macrophage M1 polarization via AKT1/SOCS1/NF-κB signaling pathway to protect against DSS-induced colitis. Int. Immunopharmacol. 2018, 57, 121–131.
- Yan, G.; Li, Y.J.; Zhao, Y.Y.; Guo, J.M.; Zhang, W.H.; Zhang, M.M.; Fua, Y.H.; Liu, Y.P. Neuroprotective Carbazole alkaloids from the stems and leaves of Clausena lenis. Nat. Prod. Res. 2021, 35, 2002–2009.
- Rastogi, M.; Ojha, R.P.; Prabu, P.C.; Parimala Devi, B.; Agrawal, A.; Dubey, G.P. Prevention of age-associated neurodegeneration and promotion of healthy brain aging in female Wistar rats by long-term use of bacosides. Biogerontology 2012, 13, 183–195.
- Son, I.S.; Kim, J.H.; Sohn, H.Y.; Son, K.H.; Kim, J.S.; Kwon, C.S. Antioxidative and hypolipidemic effects of diosgenin, a steroidal saponin of Yam (Dioscorea spp.), on high cholesterol-fed rats. Biosci. Biotechnol. Biochem. 2007, 71, 70471–70479.
- Kondeva-Burdina, M.; Krasteva, I.; Popov, G.; Manov, V. Neuroprotective and antioxidant activities of saponins’ mixture from Astragalus glycyphylloides in a model of 6-hydroxy- dopamine-induced oxidative stress on isolated rat brain synaptosomes. Pharmacia 2019, 66, 233–236.
- Abe, T.K.; Taniguchi, M. Identification of Myo-inositol hexakisphosphate (IP6) as a β- secretase 1 (BACE1) inhibitory molecule in rice grain extract and digest. FEBS Open Bio. 2014, 4, 162–167.
- Sánchez-Hernández, D.; Anderson, G.H.; Poon, A.N.; Pannia, E.; Cho, C.E.; Huot, P.S.P.; Kubant, R. Maternal fat-soluble vitamins, brain development, and regulation of feeding behavior: An overview of research. Nutr. Res. Rev. 2016, 36, 1045–1054.
- Mikkelsen, K.; Stojanovska, L.; Tangalakis, K.; Bosevski, M.; Apostolopoulos, V. Cognitive decline: A vitamin B perspective. Maturitas 2016, 93, 108–113.
- Martin, P.R.; Singleton, C.K.; Hiller-Sturmhöfel, S. The role of thiamine deficiency in alcoholic brain disease. Alcohol Res. Health 2003, 27, 134–142.
- Williams, A.; Ramsden, D. Nicotinamide: A double-edged sword. Parkinsonism Relat. Disord. 2005, 11, 413–420.
- Griffin, S.M.; Pickard, M.R.; Orme, R.P.; Hawkins, C.P.; Fricker, R.A. Nicotinamide promotes neuronal differentiation of mouse embryonic stem cells in vitro. Neuroreport 2013, 24, 1041–1046.
- Lu, L.; Tang, L.; Wei, W.; Hong, Y.; Chen, H.; Ying, W.; Chen, S. Nicotinamide mononucleotide improves energy activity and survival rate in an in vitro model of Parkinson’s disease. Exp. Ther. Med. 2014, 8, 943–950.
- Kuypers, N.J.; Hoane, M.R. Pyridoxine administration improves behavioral and anatomical outcomes after unilateral contusion injury in the rat. J. Neurotrauma 2010, 27, 1275–1282.
- Calderón-Ospina, C.A.; Nava-Mesa, M.O. B Vitamins in the nervous system: Current knowledge of the biochemical modes of action and synergies of thiamine, pyridoxine, and cobalamin. CNS Neurosci. Ther. 2020, 26, 5–13.
- Volkov, I. The master key effect of vitamin B12 in the treatment of malignancy- A potential therapy? Med. Hypotheses 2008, 70, 324–328.
- De Queiroz, K.B.; Cavalcante-Silva, V.; Lopes, F.L.; Rocha, G.A.; D’Almeida, V.; Coimbra, R.S. Vitamin B12 is neuroprotective in experimental pneumococcal meningitis through modulation of hippocampal DNA methylation. J. Neuroinflamm. 2020, 17, 1–12.
- Seitz, G.; Gerbhardt, S.; Beck, J.F.; Bohm, W.; Lode, H.N.; Niethammer, D.; Bruchelt, G. Ascorbic acid stimulates DOPA synthesis and tyrosine hydroxylase gene expression in the human neuroblastoma cell line SK-N-SH. Neurosci. Lett. 1998, 244, 33–36.
- Huang, H.O.; Caballero, B.; Chang, S.; Alberg, A.; Semba, R.; Schneyer, C.; Wilson, R.F.; Cheng, T.Y.; Prokopowicz, G.; Barnes, G.J.; et al. Multivitamin/mineral supplements and prevention of chronic disease. Evid. Rep. Technol. Assess. 2006, 139, 1–117.
- Sershen, H.; Mason, M.F.; Reith, M.E.A.; Hashim, A.; Lajtha, A. Effect of nicotine and amphetamine on the neurotoxicity of N-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) in mice. Neuropharmacology 1985, 25, 1231–1234.
- Pardo, J.V.; Pardo, P.J.; Raichle, M.E. Neural correlates of self-induced dysphoria. Am. J. Psychiatry 1993, 150, 713–719.
- Schepers, M.; Martens, N.; Tiane, A.; Vanbrabant, K.; Liu, H.B.; Lütjohann, D.; Mulder, M.; Vanmierlo, T. Edible seaweed-derived constituents: An undisclosed source of neuroprotective compounds. Neural Regen. Res. 2020, 15, 790–795.
- Javed, H.; Meeran, M.F.N.; Azimullah, S.; Adem, A.; Sadek, B.; Ojha, S.K. Plant extracts and phytochemicals targeting α-synuclein aggregation in Parkinson’s disease models. Front. Pharmacol. 2019, 9, 1–27.
- Kanhere, R.; Anjana, A.; Anbu, J.; Sumithra, M.; Ahamed, N. Neuroprotective and antioxidant potential of terpenoid fraction from Hygrophila auriculata against transient global cerebral ischemia in rats. Pharm. Biol. 2013, 51, 181–189.
- Choi, S.U.; Park, S.H.; Kim, K.H.; Choi, E.J.; Kim, S.; Park, W.K.; Zhang, Y.H.; Kim, H.S.; Jung, N.P.; Lee, C.O. The bisbenzylisoquinoline alkaloids, tetrandrine, and fangchinoline enhance the cytotoxicity of multidrug resistance-related drugs via modulation of P-glycoprotein. Anticancer Drugs 1998, 9, 255–261.
- Sun, J.; Li, M.; Zou, F.; Bai, S.; Jiang, X.; Tian, L.; Ou, S.; Jiao, R.; Bai, W. Protection of cyanidin-3-O-glucoside against acrylamide and glycidamide-induced reproductive toxicity in Leydig cells. Food Chem. Toxicol. 2018, 119, 268–274.
- Li, X.; Chu, S.; Liu, Y.; Chen, N. Neuroprotective effects of anthraquinones from Rhubarb in central nervous system diseases. Evid. Based Complement. Alternat. Med. 2019, 2019, 1–12.
- Bo, S.; Lai, J.; Lin, H.; Luo, X.; Zeng, Y.; Du, T. Purpurin, an anthraquinone that induces ROS-mediated A549 lung cancer cell apoptosis via inhibition of PI3K/AKT and proliferation. J. Pharm. Pharmacol. 2021, 73, 1101–1108.
- Liu, B.; Xie, J.; Ge, X.; Xu, P.; Wang, A.; Heb, Y.; Zhoub, Q.; Pan, L.; Chen, R. Effects of anthraquinone extract from Rheum officinale Bail on the growth performance and physio- logical responses of Macrobrachium rosenbergii under high-temperature stress. Fish Shellfish Immunol. 2010, 29, 49–57.
- Ma, L.; Zhao, L.; Hu, H.; Qin, Y.; Bian, Y.; Jiang, H.; Zhou, H.; Yu, L.; Zeng, S. Interaction of five anthraquinones from rhubarb with human organic anion transporter 1 (SLC22A6) and 3 (SLC22A8) and drug-drug interaction in rats. J. Ethnopharmacol. 2014, 153, 864–871.
- Aldbass, A.; Amina, M.; Al Musayeib, N.M.; Bhat, R.S.; Al-Rashed, S.; Marraiki, N.; Fahmy, R.; El-Ansary, A. Cytotoxic and anti-excitotoxic effects of selected plant and algal extracts using COMET and cell viability assays. Sci. Rep. 2021, 11, 8512.
- Yang, N.; Xiang, L.; Cao, F.; Li, H.; Wang, X. Research progress on the effects and mechanisms of emodin in Tumor Metastasis. Zhong Liu Yao Xue 2016, 6, 173–177.
- Gutierres, J.M.; Carvalho, F.B.; Schetinger, M.R.C.; Rodrigues, M.V.; Schmatz, R.; Pimentel, V.C.; Vieira, J.M.; Rosa, M.M.; Marisco, P.; Ribeiro, D.A.; et al. Protective effects of anthocyanins on the ectonucleotidase activity in the impairment of memory induced by scopolamine in adult rats. Life Sci. 2012, 91, 1221–1228.
- Jeter, C.B.; Hergenroeder, G.W.; Ward, N.H., III; Moore, A.N.; Dash, P.K. L-arginine, and its metabolite levels: A possible link to cerebral blood flow, extracellular matrix remodeling, and energy status. J. Neurotrauma 2012, 29, 119–127.
- Liu, H.; Goodman, J.C.; Robertson, C.S. The effects of L-arginine on cerebral hemodynamics after controlled cortical impact injury in the mouse. J. Neurotrauma 2004, 19, 327–334.
- Cherian, L.; Chacko, G.; Goodman, C.; Robertson, C.S. Neuroprotective effects of L-arginine administration after cortical impact injury in rats: Dose-response and time window. J. Pharmacol. Exp. Ther. 2003, 304, 617–623.
- Bitner, B.R.; Brink, D.C.; Mathew, L.C.; Pautler, R.G.; Robertson, C.S. Impact of arginase II on CBF in experimental cortical impact injury in mice using MRI. J. Cereb. Blood Flow Metab. 2010, 30, 1105–1109.
- De George, M.L.; Marlowe, D.; Werner, E.; Soderstrom, K.E.; Stock, M.; Mueller, A.; Bohn, M.C.; Kozlowski, D.A. Combining glial cell line-derived neurotrophic factor gene delivery (AdGDNF) with L-arginine decreases contusion size but not behavioral deficits after traumatic brain injury. Brain Res. 2011, 1403, 45–56.
- Meloni, B.P.; Craig, A.J.; Milech, N.; Hopkins, R.M.; Watt, P.M.; Knuckey, N.W. The neuroprotective efficacy of cell-penetrating peptides TAT, penetratin, Arg-9, and Pep-1 in glutamic acid, kainic acid, and in vitro ischemia injury models using primary cortical neuronal cultures. Cell. Mol. Neurobiol. 2014, 34, 173–181.
- Meloni, B.P.; Milani, D.; Cross, J.L.; Clark, V.W.; Edwards, A.B.; Anderton, R.S.; Blacker, D.J.; Knuckey, N.W. Assessment of the neuroprotective effects of arginine-rich protamine peptides, poly-arginine peptides (R12-Cyclic, R22), and arginine-tryptophan-containing peptides following in vitro excitotoxicity and/or permanent middle cerebral artery occlusion in rats. Neuromolecular Med. 2017, 19, 271–285.
- Edwards, A.B.; Cross, J.L.; Anderton, R.S.; Knuckey, N.W.; Meloni, B.P. Poly-arginine R18 and R18D (D-enantiomer) peptides reduce infarct volume and improve behavioral outcomes following perinatal hypoxic-ischaemic encephalopathy in the P7 rat. Mol. Brain 2018, 11, 1–12.
- Sachdeva, A.K.; Chopra, K. Lycopene abrogates Aβ(1-42)-mediated neuroinflammatory cascade in an experimental model of Alzheimer’s disease. J. Nutr. Biochem. 2015, 26, 736–744.
- Lorenzo, Y.; Azqueta, A.; Luna, L.; Bonilla, F.; Domínguez, G.; Collins, A.R. The carotenoid β-cryptoxanthin stimulates the repair of DNA oxidation damage in addition to acting as an antioxidant in human cells. Carcinogenesis 2008, 30, 308–314.
- Brimson, J.M.; Brimson, S.J.; Brimson, C.A.; Rakkhitawatthana, V.; Tencomnao, T. Rhinacanthus nasutus extracts prevent glutamate and amyloid-β neurotoxicity in HT-22 mouse hippocampal cells: Possible active compounds include lupeol, stigmasterol, and β-sitosterol. Int. J. Mol. Sci. 2012, 13, 5074–5097.
- Hamedi, A.; Ghanbari, A.; Saeidi, V.; Razavipour, R.; Azari, H. Effects of β-sitosterol oral administration on the proliferation and differentiation of neural stem cells. J. Funct. Foods 2014, 8, 252–258.
- Hamedi, A.; Ghanbari, A.; Razavipour, R.; Saeidi, V.; Zarshenas, M.M.; Sohrabpour, M.; Azari, H. Alyssum homolocarpum seeds: Phytochemical analysis and effects of the seed oil on neural stem cell proliferation and differentiation. J. Nat. Med. 2015, 69, 387–396.
- Cheng, F.C.; Jinn, T.R.; Hou, R.C.W.; Tzen, J.T.C. Neuroprotective effects of sesamin and sesamolin on Gerbil brain in cerebral ischemia. Int. J. Biomed. 2006, 2, 284–288.
- Shimmyo, Y.; Kihara, T.; Akaike, A.; Niidome, T.; Sugimoto, H. Multifunction of myricetin on A beta: Neuroprotection via a conformational change of A-beta and reduction of A beta via the interference of secretases. J. Neurosci. Res. 2008, 86, 368–377.
- Jones, J.R.; Lebar, M.D.; Jinwal, U.K.; Abisambra, J.K.; Koren, J., III; Blair, L.; O’Leary, J.C.; Davey, Z.; Trotter, J.; Johnson, A.G.; et al. The diarylheptanoid (+)-aR, 11S-myricanol, and two flavones from bayberry (Myrica cerifera) destabilize the microtubule-associated protein Tau. J. Nat. Prod. 2011, 74, 38–44.
- Pinto, N.B.; Alexandre, B.; Neves, K.R.T.; Silva, A.H.; Leal, L.K.A.M.; Viana, G.S.B. Neuroprotective properties of the standardized extract from Camellia sinensis (Green Tea) and its main bioactive components, epicatechin and epigallocatechin gallate, in the 6-OHDA model of Parkinson’s disease. Evid. Based Complement. Alternat. Med. 2015, 2015, 161092.
- Cifuentes, J.; Salazar, V.A.; Cuellar, M.; Castellanos, M.C.; Rodríguez, J.; Cruz, J.C.; Muñoz-Camargo, C. Antioxidant and Neuroprotective Properties of Non-Centrifugal Cane Sugar and Other Sugarcane Derivatives in an In Vitro Induced Parkinson’s Model. Antioxidants 2021, 10, 1040.
- Mendonça-Junior, F.J.B.; Scotti, M.T.; Muratov, E.N.; Scotti, L.; Nayarisseri, A. Natural Bioactive Products with Antioxidant Properties Useful in Neurodegenerative Diseases 2020. Oxid. Med. Cell. Longev. 2021, 2021, 1–2.
- Hossain, M.T.; Furhatun-Noor; Asadujjaman; Matin, A.; Tabassum, F.; Ar Rashid, H. A Review study on the pharmacological effects and mechanism of action of Tannins. Eur. J. Pharm. Med. Res. 2021, 8, 5–10.
- Soldado, D.; Bessa, R.J.B.; Jeronimo, E. Condensed Tannins as Antioxidants in Ruminants—Effectiveness and Action Mechanisms to Improve Animal Antioxidant Status and Oxidative Stability of Products. Animals 2021, 11, 3243.
- Ali, T.; Kim, T.; Rehman, S.U.; Khan, M.S.; Amin, F.U.; Khan, M.; Ikram, M.; Kim, M.O. Natural Dietary Supplementation of Anthocyanins via PI3K/Akt/Nrf2/HO-1 Pathways Mitigate Oxidative Stress, Neurodegeneration, and Memory Impairment in a Mouse Model of Alzheimer’s Disease. Mol. Neurobiol. 2018, 55, 6076–6093.
- Puiggròs, F.; Llópiz, N.; Ardévol, A.; Bladé, C.; Arola, L.; Salvadó, M.J. Grape Seed Procyanidins Prevent Oxidative Injury by Modulating the Expression of Antioxidant Enzyme Systems. J. Agric. Food Chem. 2005, 53, 6080–6086.
- Echeverry, C.; Arredondo, F.; Abin-Carriquiry, J.A.; Midiwo, J.O.; Ochieng, C.; Kerubo, L.; Dajas, F. Pretreatment with Natural Flavones and Neuronal Cell Survival after Oxidative Stress: A Structure-Activity Relationship Study. J. Agric. Food Chem. 2010, 58, 2111–2115.
- Boesch-Saadatmandi, C.; Pospissil, R.T.; Graeser, A.; Canali, R.; Boomgaarden, I.; Doering, F.; Wolffram, S.; Egert, S.; Mueller, M.J.; Rimbach, G. Effect of Quercetin on Paraoxonase 2 Levels in RAW264.7 Macrophages and in Human Monocytes- Role of Quercetin Metabolism. Int. J. Mol. Sci. 2009, 10, 4168–4177.
- Costa, L.G.; Tait, L.; de Laat, R.; Dao, K.; Giordano, G.; Pellacani, C.; Cole, T.B.; Furlong, C.E. Modulation of paraoxonase 2 (PON2) in mouse brain by the polyphenol quercetin: A mechanism of neuroprotection? Neurochem. Res. 2014, 38, 1809–1818.
- Grewal, A.K.; Singha, T.G.; Sharma, D.; Sharma, V.; Singh, M.; Rahman, H.; Najda, A.; Walasek-Janusz, M.; Kamel, M.; Albadrani, G.M.; et al. Mechanistic insights and perspectives involved in neuroprotective action of quercetin. Biomed. Pharmacother. 2021, 140, 111729.
- Zhang, Z.J.; Cheang, L.C.V.; Wang, M.W.; Lee, S.M. Quercetin exerts a neuroprotective effect through inhibition of the iNOS/NO system and pro-inflammation gene expression in PC12 cells and in zebrafish. Int. J. Mol. Med. 2011, 27, 195–203.
- Ansari, M.A.; Abdul, H.M.; Joshi, G.; Opii, W.O.; Butterfield, D.A. Protective effect of quercetin in primary neurons against Abeta(1-42): Relevance to Alzheimer’s disease. J. Nutr. Biochem. 2009, 20, 269–275.
- Salehi, B.; Machin, L.; Monzote, L.; Sharifi-Rad, J.; Ezzat, S.M.; Salem, M.A.; Merghany, R.M.; El Mahdy, N.M.; Kılıç, C.S.; Sytar, O.; et al. Therapeutic Potential of Quercetin: New Insights and Perspectives for Human Health. ACS Omega 2020, 5, 11849–11872.
- Khazdair, M.R.; Ghafari, S.; Sadeghi, M. Possible therapeutic effects of Nigella sativa and its thymoquinone on COVID-19. Pharm. Biol. 2021, 59, 696–703.
- Wang, J.; Fang, X.; Ge, L.; Cao, F.; Zhao, L.; Wang, Z.; Xiao, W. Antitumor, antioxidant and anti-inflammatory activities of kaempferol and its corresponding glycosides and the enzymatic preparation of kaempferol. PLoS ONE 2018, 13, e0197563.
- Kavya Bhatt, S.; Manjunatha Javagal, R.; Shashirekha Nanjarajurs, M.; Eligar, S.M. In vitro anti-inflammatory property of a Quercetin-3-O-diglucoside-7-O-glucoside characterized from fresh leaves of Trigonella foenum-graecum L. Int. J. Food Prop. 2021, 24, 1438–1452.
- Isah, T. Anticancer Alkaloids from Trees: Development into Drugs. Pharmacognosy Reviews. Pharmacogn. Rev. 2016, 10, 90–99.
- Huang, Y.; Cui, S.; Cui, X.; Cao, Q.; Ding, H.; Song, J.; Hu, X.; Ye, H.; Yu, B.; Sheng, Z.; et al. Tetrandrine, an alkaloid from S. tetrandra exhibits anti-hypertensive and sleep-enhancing effects in SHR via different mechanisms. Phytomedicine 2016, 23, 1821–1829.
- Yang, Z.; Zhang, D.; Ren, J.; Yang, M. Skimmianine, a furoquinoline alkaloid from Zanthoxylum nitidum as a potential acetylcholinesterase inhibitor. Med. Chem. Res. 2012, 2, 722–725.
- Shimizu, M.; Shirakami, Y.; Moriwaki, H. Targeting Receptor Tyrosine Kinases for Chemoprevention by Green Tea Catechin, EGCG. Int. J. Mol. Sci. 2008, 9, 1034–1049.
- Coșarcă, S.; Tanase, C.; Muntean, D.L. Therapeutic aspects of Catechin and its derivatives—An Update. Acta Biol. Marisiensis 2019, 2, 21–29.
- Yang, W.; Chen, X.; Li, Y.; Guo, S.; Wang, Z.; Yu, X. Advances in Pharmacological Activities of Terpenoids. Nat. Prod. Commun. 2020, 15, 1–13.
- Yoo, K.; Park, S. Terpenoids as potential anti-Alzheimer’s disease therapeutics. Molecules 2012, 17, 3524–3538.
- Kwon, K.H.; Murakami, A.; Tanaka, T.; Ohigashi, H. Dietary rutin, but not its aglycone quercetin, ameliorates dextran sulfate sodium-induced experimental colitis in mice: Attenuation of pro-inflammatory gene expression. Biochem. Pharmacol. 2005, 69, 395–406.
- Shamim, A.; Mahmood, T.; Ahsan, F.; Kumar, A.; Bagga, P. Lipids: An insight into the neurodegenerative disorders. Clin. Nutr. Exp. 2018, 20, 1–19.