Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Egg protein is a remarkably abundant source of protein, with an amino acid score of 100 and the highest net protein utilization rate. However, there have been relatively fewer studies investigating the health benefits of egg protein. The available information regarding the health benefits of egg proteins based on human studies has been summarized. In particular, studies conducted on the characteristics of egg whites, as they are high in pure protein, have reported their various health functions, such as increases in muscle mass and strength enhancement, lowering of cholesterol, and visceral fat reduction.
- egg
- protein
- egg protein hydrolysate
- egg white
- cholesterol-lowering
- visceral fat-lowering
- antifatigue
1. Introduction
Chicken eggs are a highly nutritious food, which contain most of the essential nutrients for human life except for vitamin C and dietary fiber [1]. However, as they also contain cholesterol, patients with dyslipidemia may be advised to avoid their consumption. In healthy individuals, eggs have been reported to have no effect on blood lipid content [2]. This is because even if cholesterol is ingested from the diet, the body has mechanisms by which to control its levels by reducing cholesterol synthesis in the liver [3].
The various health benefits of eggs have been demonstrated in the existing literature, such as the improvement of blood lipids due to the intake of the phospholipids found in the egg yolk, the improvement of cognitive function due to the choline in the egg yolk [4][5], and increased bone density due to the calcium derived from the eggshells [6]. In addition, egg whites are characterized by their high dry weight protein content. Previous studies have described the nutritional benefits of protein using whey and soy proteins. In contrast, egg-derived protein research has generally been focused on allergens. However, in recent years, there have been an increasing number of reports describing the nutritional benefits of egg protein.
The amino acid compositions of egg white protein (EWP; dried egg white), casein, and soy protein isolate are shown in Table 1 [7]. One characteristic of EWP is that it contains approximately the same amount of branched-chain amino acids as milk protein. It also has more sulfur-containing amino acids than other protein sources. The amino acid score for EWP is 100, and its net protein utilization is reported to be higher than that of whey protein [8]. Due to the high quality of EWP, in animal experiments, the consumption of EWP was found to increase body protein [9]. Furthermore, body fat was found to decrease, indicating that EWP is expected to affect health by positively impacting metabolic syndrome. Proteins are also present in eggshell membranes, and these have been the subject of various functional studies [10][11]. In this entry, researchers will introduce the health benefits of egg proteins from a nutritional perspective, with a focus on the existing evidence for EWP, which has been reported in humans.
Table 1. Amino acid compositions of casein, dried egg white, and soy protein isolates (g/100 g) [7].
| Casein | Dried Egg White | Soy Protein Isolate | |
|---|---|---|---|
| Ile | 5.0 | 4.4 | 4.0 |
| Leu | 8.5 | 7.3 | 7.0 |
| Lys | 7.2 | 6.1 | 5.5 |
| Met | 2.7 | 3.2 | 1.1 |
| Cys | 0.4 | 2.5 | 1.1 |
| Phe | 4.6 | 5.1 | 4.6 |
| Tyr | 5.2 | 3.9 | 3.5 |
| Thr | 4.0 | 4.0 | 3.7 |
| Trp | 1.1 | 1.3 | 1.2 |
| Val | 6.2 | 5.8 | 4.1 |
| His | 2,7 | 2.1 | 2.4 |
| Arg | 3.4 | 5.0 | 6.9 |
| Ala | 2.7 | 5.3 | 3.6 |
| Asp | 6.3 | 9.3 | 10.0 |
| Gul | 19.0 | 12.0 | 17.0 |
| Gly | 1.7 | 3.2 | 3.6 |
| Pro | 10.0 | 3.3 | 4.7 |
| Ser | 5.2 | 6.0 | 5.1 |
Basic research, such as animal studies and component analysis, has been the primary method of research to determine the health benefits of egg proteins. The health benefits of egg protein hydrolysate and ovotransferrin have been reviewed [12][13][14][15], although based on the results of basic research. In one review considering human studies, the health benefits of the egg itself have been reported [16][17]. Alternatively, a review summarizing the efficacy of egg protein in muscle strengthening, the prevention of sarcopenia, and the prevention of metabolic syndrome of EWR is also available [18][19]. To harness the health value of eggs at the societal level, it is important to highlight its effectiveness in human studies, identify the mechanisms of the active ingredients, and to make them easily accessible for consumption as foods and pharmaceuticals. However, review articles covering these contents have not been reported.
In addition, it is not comfortable to consume EWP as is, especially egg whites and dried egg whites, due to their physical properties and flavor. Consequently, lactic-fermented egg whites (LE) were developed, which can be consumed without additives by improving the flavor through the lactic acid fermentation of egg whites [20]. LE have almost the same electrophoretic band positions and pepsin degradability as unheated egg whites (Figure 1) [21], making it possible to verify the effects of the EWP in humans using LE. In this entry, researchers summarize the health benefits of egg protein and report on its effectiveness in humans and its mechanisms.
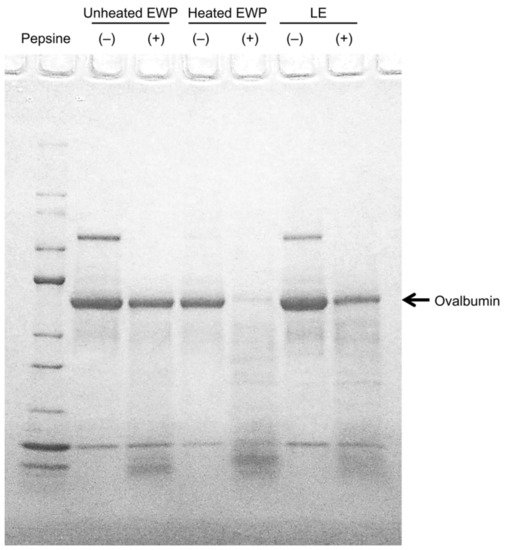
Figure 1. Electrophoresis of unheated EWP, heated EWP, LE, and its pepsin hydrolysates. Note: (+) protein pepsin hydrolysate; (−) protein before acting with pepsin [21].
2. Muscle Mass and Strength-Enhancing Effects of Egg White Protein in Combination with Exercise
EWP reportedly has protein digestibility corrected amino acid scores (PDCAAS) and digestible indispensable amino acid scores (DIAAS) that are comparable to those of whey protein [22]. In addition, the net protein utilization rate for unheated, half-boiled, and heated states of EWP is reportedly higher than that of whey protein, which is known as a good protein source [8]. In animal studies, the body protein level and gastrocnemius muscle weight in rats fed EWP were found to be significantly increased when compared to those fed casein [9], making it a promising protein supplement for muscle maintenance and building.
Suzuki et al. reported that in addition to mild resistance exercise, 15 g of EWP and 18 g of sugar per day for five weeks increased the isometric muscle strength in the forearm flexors and legs when compared to pre-consumption [23].
When women over 50 years of age were given lactic-fermented egg white (8 g per day as EWP) for eight weeks in addition to twice-weekly fitness club exercise, it was observed that the skeletal muscle weight of the limbs increased significantly in women over 55 years of age when the lactic-fermented egg white intake group were compared to the control group. Furthermore, in these women, the isometric knee extension muscle strength was increased when compared to that before ingestion [24]. This indicates that the ingestion of EWP during exercise increases both the muscle mass and strength.
There are reports, however, that the muscle-mass- and muscle-strength-enhancing effects are stronger for whole egg protein than for EWP, and future studies are expected to resolve this [25][26].
In studies using egg white as a protein supplement, it is not appropriate to use liquid egg white itself due to its unfavorable taste and flavor. Therefore, dried egg white is likely to be used. However, there are numerous issues regarding the flavor and physical properties of dried egg whites when consumed without additives. Consequently, the development of dried egg whites that are easy to consume or the use of LE is desirable.
3. Egg White Protein Improves Lipid Metabolism
3.1. Visceral Fat Reduction Effects
EWP is classified as a high-quality protein source, and when fed to rats their whole-body protein content was found to be higher when compared to those fed casein. The EWP intake was, thus, shown to increase body protein and decrease total body fat levels [9]. In contrast, when fed the EWP, the triglyceride and leptin concentrations in the rat tail vein plasma were found to be significantly lower in comparison with those fed casein, indicating that EWP could reduce visceral fat levels [9]. Consequently, the effects of EWP on the visceral fat in rats was further investigated and compared with casein. The results indicated that the EWP could significantly reduce white adipose tissue weight and white adipocyte area levels. EWP consumption was also found to result in significantly higher gastrocnemius muscle weight in comparison with casein [9]. This indicates that egg white is a good source of protein that can be utilized to increase body protein, i.e., muscle mass. Since muscle tissue is the primary site of fat oxidation [27], it was hypothesized that the consumption of egg whites may reduce both body fat and visceral fat levels.
Unheated egg whites have been used in previous studies, and it was reported that the net protein utilization rates of EWP are comparable between unheated and heated egg whites [7]. Consequently, it was hypothesized that heated egg whites would reduce visceral fat levels in a similar manner to unheated egg whites. The effects of heated egg whites (boiled egg: 95 °C for 10 min) on visceral fat were, thus, compared to unheated egg whites and LE, with casein being used as a negative control. Although the unheated egg whites and LE reduced the visceral fat levels when compared to casein, the heated egg whites did not [20]. These results indicate that there may be mechanisms other than the provided hypothesis, namely that egg whites reduce visceral fat levels by increasing muscle mass. A summary of the known visceral fat reduction mechanisms of EWP is provided in Figure 2.
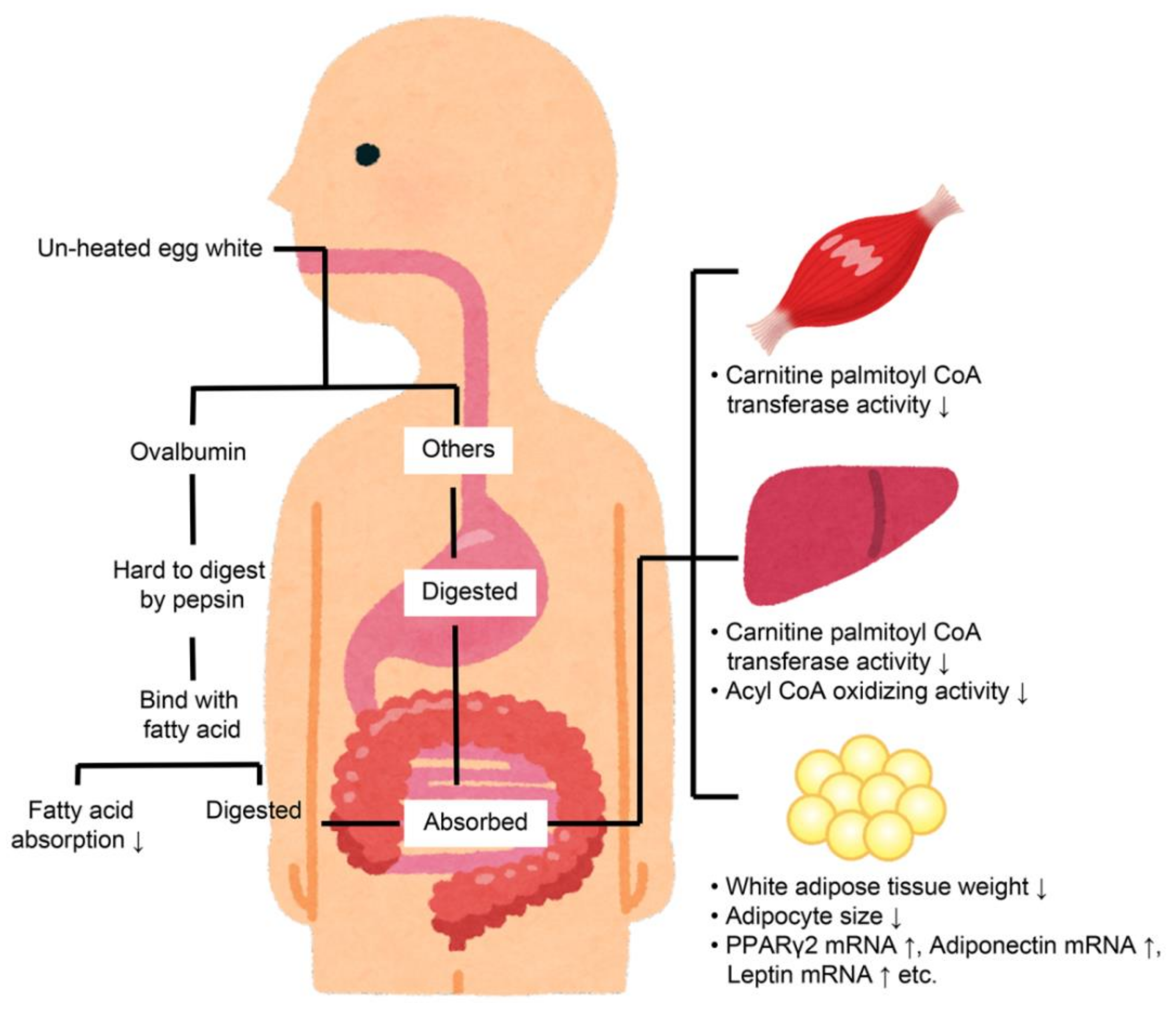
An allergy study found that the ovalbumin in EWP is not easily degraded by pepsin in unheated egg whites but is easily degraded in heated egg whites [31]. Therefore, if ovalbumin in LE is less likely to be degraded by pepsin, the digestibility of pepsin may be involved in the visceral fat-reducing mechanism. Therefore, when researchers evaluated the pepsin degradability of ovalbumin in LE, it was found that the ovalbumin in LE was not easily degraded by pepsin [21].
Pepsin is a gastric digestive enzyme, indicating that the main factor in the visceral fat-reducing effect of EWP may be occurring in the gastrointestinal tract. The triglyceride absorption into the lymph of rats undergoing thoracic lymph duct cannulation surgery was also reduced [28]. It was shown that the suppression of lipid absorption by ovalbumin in unheated egg whites may have reduced the visceral fat level. In an in vitro study by Handa et al. (1999), ovalbumin was reported to bind to fatty acids [32]. This indicated that ovalbumin, which is not easily affected by pepsin, can bind to fatty acids degraded by lipase in the digestive tract, thereby suppressing fatty acid absorption.
However, while the suppression of lipid absorption by EWP is approximately 20% higher than that by casein [28], the visceral fat reduction effect is 30% less than that of casein [9]. Despite the differences in experimental conditions, these tests were conducted by feeding diets containing 20% casein or EWP, and the visceral fat reductions by EWP could not be explained solely by the suppression of lipid absorption. Other mechanisms for reducing visceral fat using EWP include increased β-oxidation in the muscle and liver, as well as a reduction in the visceral adipocyte area [9].
The enhancement of β-oxidation in the liver was reported in a study in which rats were fed diets containing 20% casein or EWP. The results showed that EWP consumption resulted in significantly higher activity levels for carnitine palmitoyltransferase and acyl-CoA oxidase, which are involved in β-oxidation in the liver, when compared to casein [9].
The enhancement of β-oxidation in the muscle was identified in a similar study in which rats were a fed diet containing 20% casein or EWP, and the EWP diet was found to increase gastrocnemius muscle mass and showed higher levels of enzyme activities related to β-oxidation in the muscle when compared to casein [33].
Studies investigating the decrease in visceral fat area in rats have also shown an increase in the expression of a series of genes, including peroxisome proliferator-responsive receptor (PPAR) γ2 and adiponectin in visceral adipocytes, as well as improved insulin sensitivity [29][30]. The reductions in visceral fat by EWPs were also thought to be caused by these effects.
The following two possibilities were considered as explanations for the increased β-oxidation in the muscle and liver and the increased gene expression in the visceral adipocytes. The first was the possibility of a secondary effect for lipid absorption suppression. The second was the possibility of a direct effect of the EWP-derived components on the liver, muscle, and adipose tissue. For the latter, the absorption rate of the unheated EWP was reported to be over 90% [21]. In addition, EWP was digested into peptides in the gastrointestinal tract [34]. Therefore, the absorption of peptides derived from EWP may generate these effects. In the liver, soy-derived peptides reportedly enhance the gene expression of enzymes related to β-oxidation in a mouse model for diabetes [35]. For white fat cells, it has been reported that soy hydrolysate enhances the gene expression of PPARγ2, albeit this has only been found for in vitro results [36]. Based on these findings, researchers expected to find a peptide with similar effects in the EWP. The identification of the peptide sequence that exerts this effect in EWP will be a subject for future studies.
Reductions in visceral fat using EWP in humans have not yet been investigated. Like EWP, lactoferrin and β-conglycinin from soybeans have also been reported to reduce visceral fat [37][38]. Lactoferrin is reported to reduce visceral fat at 300 mg daily and β-conglycinin at 5 g per day [37][38]. The inhibition of lipid absorption has been reported as a mechanism for visceral fat reduction using β-conglycinin [39], and the inhibition of lipid absorption by ovalbumin, ovotransferrin, and lysozyme has been reported as a visceral fat reduction mechanism for EWP [28][40]. To consume 5 g of these three components in equal measure, 8 g of LE as EWP needs to be consumed.
At the same time, the function of ovotransferrin in EWP was found to be similar to that of lactoferrin. If ovotransferrin has the same physiological activity as lactoferrin, it was hypothesized that consuming 3 g of lactic-fermented egg white per day as protein would reduce the level of visceral fat. Based on this, the minimum effective amount of lactic-fermented egg white was determined to be 3–8 g of protein per day.
Therefore, the effects of LE containing 6 or 8 g of EWP per day or 8 g of whey protein per day was evaluated on the visceral fat levels of 22 Japanese adult males (overall) with a BMI ≥ 24 and waist ≥ 85 cm for eight weeks. The results showed that the consumption of lactic-fermented egg white, which was equivalent to 8 g of EWP per day, reduced the visceral fat area when compared to that before consumption. A further analysis in subjects with a BMI > 25 showed that 8 g per day of lactic-fermented egg white as EWP significantly reduced the visceral fat area and visceral fat/subcutaneous fat ratio when compared to pre-consumption levels or when whey protein was used instead [21]. The results indicate that 8 g of EWP per day may reduce the total visceral fat area.
4. Antifatigue Effects of Egg White Protein Hydrolysate
The physiological functions of EWP have been described. However, various health effects of hydrolyzed proteins, which are artificially prepared using enzymes, especially peptides, derived from milk and soybeans have also been reported [41][42][43]. The physiological functions of egg white protein hydrolysate (EWH: mainly Peptifine®, Kewpie Corporation, Tokyo, Japan) were, thus, assessed.
The absorption and nutritional value of whey protein hydrolysate with a molecular weight of 500 and EWH with a molecular weight of 2500 were assessed (Egg White Peptide EP-3, Henningsen Foods Inc., Omaha, NE, USA). The results showed that for the EWH, the absorption rate of amino acids into the portal vein was faster than with the whey protein hydrolysate, despite its larger molecular weight. The PDCAAS and DIAAS of egg white hydrolysate as well as whey protein and its hydrolysate. In addition, the net protein utilization was significantly higher for EWH than whey protein hydrolysate [22].
One of the ongoing challenges for EWP is its bitter flavor. To address this, a new hydrolysate that was less bitter was developed by enzymatically degrading egg whites and collecting the soluble fraction (Peptifine®) [44].
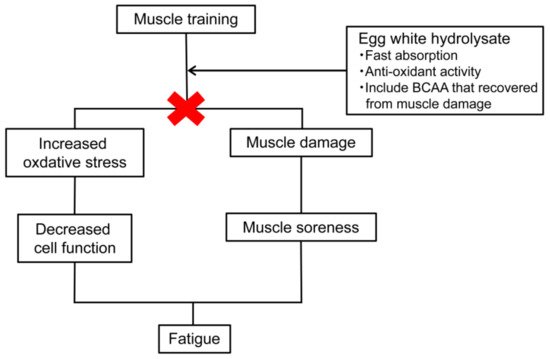
Egg whites have high amounts of sulfur-containing amino acids and branched-chain amino acids (BCAA) [45]. Since sulfur-containing amino acids are the source of glutathione [46], they are expected to have antioxidant properties. Antioxidant effects are known to prevent various diseases, including atherosclerotic diseases [47], and have also been reported to lead to antifatigue effects. In addition, BCAAs have been reported to improve muscle fatigue [48], and since the blood BCAA concentration increases when EWP is ingested and decreases after exercise [49], an antifatigue effect of the BCAAs can also be expected (Figure 3).
No antifatigue effects have been reported for proteins, but they have been reported for peptides. Imidazole peptides derived from chickens reportedly have antioxidant properties, and consequently antifatigue effects [50]. Peptides from whey have been reported to increase swimming times in forced swimming mice by chelating radical scavengers and iron [51].
EWH has also been reported to have antioxidant properties [52]. In addition, the pepsin degradation products of egg whites have also been reported to have in vivo antioxidant and antifatigue effects [53].
The effects of EWH (Peptifine®) on the swimming times of weight-loaded forced swimming mice were consequently investigated. Seven-week-old male ddY mice were divided into a casein group, an EWH group, and an EWP group and fed diets containing each for 14 days. The swimming times were evaluated daily starting on the 11th day of the study. On the last day of the test, blood samples were taken for a blood test. The results showed that the swimming time of the EWH group was significantly longer than for the casein and EWP groups on the 14th day of the testing. The reason for this was a decrease in the concentration of hexanoyl lysine in the blood of the EWH group. In summary, the egg white hydrolysate was found to prolong the swimming time in forced swimming, and the mechanism of the prolonged swimming time was thought to be due to the antifatigue effect of its antioxidant action (Figure 3) [54].
In healthy marathon runners, 7.5 g of EWH (Peptifine®) per day for eight weeks was found to improve the subjective symptoms of physical and overall fatigue when compared to the controls. Additionally, at this time, serum aspartate aminotransferase (AST), alanine aminotransferase (ALT), and lactate dehydrogenase were found to be significantly lower than in the controls at week 8 [55].
Similarly, when marathon runners were given 2.5 g and 5 g of EWH per day for eight weeks, physical fatigue was significantly lower in the group given 5 g of EWH per day when compared to the control group [55].
These results indicate that the minimum effective dose of EWH to exert its antifatigue effect is 5 g. In addition, when athletes were continuously given 5 g of EWH (Peptifine®) per day for two weeks, the psychological fatigue was reduced when compared to the control [56]. It was, thus, determined that EWH could be used effectively to improve the exercising capacity of humans, as it is a good source of protein and raw materials for muscles due to its amino acid composition, and it is quickly absorbed and improves physical fatigue through its antioxidant effects.
This entry is adapted from the peer-reviewed paper 10.3390/foods11152309
References
- Ruxto, C. Recommendation for the use of egg in the diet. Nurs. Stand. 2010, 24, 47–55.
- Sugano, M.; Matsuoka, R. Nutritional viewpoint on egg and cholesterol. Food 2021, 10, 494.
- Ezaki, O.; Sakono, M.; Miyake, M.; Mito, N. Concepts for Reference Intake of Cholesterol in the Japanese Population. J. Jpn. Soc. Nutr. Food Sci. 2005, 58, 69–83. (In Japanese)
- Masuda, Y.; Kokubu, T.; Yamashita, M.; Ikeda, H.; Inoue, S. Egg phosphatidylcholine combined with vitamin B12 improved memory impairment following lesioning nucleus basalis in rats. Life Sci. 1998, 62, 813–822.
- Yin, M.; Matsuoka, R.; Xi, Y.; Wang, X. Comparison of egg yolk and soybean phospholipids on hepatic fatty acid profile and liver protection in rats fed a high fructose diets. Foods 2021, 10, 1569.
- Sakai, S.; Hien, V.T.T.; Tuyen, L.D.; Duc, H.A.; Masuda, Y.; Yamamoto, S. Effects of Eggshell Calcium Supplementation on Bone Mass in Postmenopausal Vietnamese Women. J. Nutr. Sci. Vitaminol. 2017, 63, 120–124.
- Ministry of Education, Culture, Sports, Science and Technology, Standard Tables of Food Composition in Japan (Seventh Revised Version). 2015; Available online: https://www.mext.go.jp/en/policy/science_technology/policy/title01/detail01/1374030.htm (accessed on 9 June 2022).
- Matsuoka, R.; Takahashi, Y.; Kimura, M.; Masuda, Y.; Kunou, M. Heating has no effect on the net protein utilisation from egg whites in rats. Sci. World J. 2017, 2017, 6817196.
- Matsuoka, R.; Shirouchi, B.; Umegatani, M.; Fukuda, M.; Muto, A.; Masuda, Y.; Kunou, M.; Sato, M. Dietary egg white protein increases body protein mass and reduces body fat mass through an acceleration of hepatic β-oxidation in rats. Br. J. Nutr. 2017, 118, 423–430.
- Yoo, J.H.; Kim, J.K.; Yang, H.J.; Park, K.M. Egg shell membrane hydrolysates on the UVB-radiation-induced wrinkle formation in SKH-1 hairless mice. Korean J. Food. Sci. 2015, 35, 58–70.
- Wedekind, K.J.; Ruff, K.J.; Atwell, C.A.; Evans, J.L.; Bendele, A.M. Beneficial effects of natural eggshell membrane (NEM) on m-ultiple indices of arthritis in collagen-induced arthritic rats. Mod. Rheumatol. 2017, 27, 838–848.
- Liao, W.; Jahandideh, F.; Fan, H.; Son, M.; Wu, J. Egg Protein-Derived Bioactive Peptides: Preparation, Efficacy, and Absorption. Adv. Food Nutr. Res. 2018, 85, 1–58.
- Legros, J.; Jan, S.; Bonnassie, S.; Gautier, M.; Croguennec, T.; Pezennec, S.; Cochet, M.F.; Nau, F.; Andrews, S.C.; Baron, F. The Role of Ovotransferrin in Egg-White Antimicrobial Activity: A Review. Foods 2021, 10, 823.
- Lee, J.H.; Paik, H.D. Anticancer and immunomodulatory activity of egg proteins and peptides: A review. Poult. Sci. 2019, 98, 6505–6516.
- Mine, Y. Egg proteins and peptides in human health--chemistry, bioactivity and production. Curr. Pharm. Des. 2007, 13, 875–884.
- Sanlier, N.; Üstün, D. Egg consumption and health effects: A narrative review. J. Food Sci. 2021, 86, 4250–4261.
- Marventano, S.; Godos, J.; Tieri, M.; Ghelfi, F.; Titta, L.; Lafranconi, A.; Gambera, A.; Alonzo, E.; Sciacca, S.; Buscemi, S.; et al. Egg consumption and human health: An umbrella review of observational studies. Int. J. Food Sci. Nutr. 2020, 71, 325–331.
- Shirouchi, B.; Matsuoka, R. Alleviation of Metabolic Syndrome with Dietary Egg White Protein. J. Oleo Sci. 2019, 68, 517–524.
- Puglist, M.J.; Luz Fernandez, M. The Health Benefits of Egg Protein. Nutrients 2022, 14, 2904.
- Arimitsu, K.; Kojima, T.; Matsuoka, R.; Narita, K.; Idei, A. The good function of lactic fermented egg white “Lacty Egg”. Jpn. J. Food. Eng. 2015, 16, 79–82. (In Japanese)
- Matsuoka, R.; Takahashi, Y.; Muto, A.; Kimura, M. Heated egg white has no effect, but lactic fermented and unheated egg white reduces abdominal fat in rats. Lipids Health Dis. 2019, 18, 187.
- Matsuoka, R.; Kurihara, H.; Nishijima, N.; Oda, Y.; Handa, A. Egg white hydrolysate retains the nutritional values and is quickly absorbed in rats. Sci. World J. 2019, 2019, 5475302.
- Kato, Y.; Sawada, A.; Numao, S.; Suzuki, M. Chronic effects of light resistance exercise after ingestion of a high-protein snack on increase of skeletal muscle mass and strength in young adults. J. Nutr. Sci. Vitaminol. 2011, 57, 233–238.
- Takanami, Y.; Kawai, Y.; Nakata, C.; Nishiyama, H.; Matsuoka, R. The combined effect of excise and lactic fermented egg white on the skeletal muscle function and muscle mass in middle-aged and elderly women. In Proceedings of the 70th Annual Meeting of the Japanese Society of Physical Fitness and Sports Medicine, Wakayama, Japan, 18–20 September 2015; Tajima, H., Ed.; The Japanese Society of Physical Fitness and Sports Medicine: Tokyo, Japan, 2015.
- Van Vliet, S.; Shy, E.L.; Abou Sawan, S.; Beals, J.W.; West, D.W.; Skinner, S.K.; Ulanov, A.V.; Li, Z.; Paluska, S.A.; Parsons, C.M.; et al. Consumption of whole eggs promotes greater stimulation of postexercise muscle protein synthesis than consumption of isonitrogenous amounts of egg whites in young men. Am. J. Clin. Nutr. 2017, 106, 1401–1412.
- Bagheri, R.; Hooshmand Moghadam, B.; Ashtary-Larky, D.; Forbes, S.C.; Candow, D.G.; Galpin, A.J.; Eskandari, M.; Kreider, R.B.; Wong, A. Whole Egg vs. Egg White Ingestion during 12 weeks of Resistance Training in Trained Young Males: A Randomized Controlled Trial. J. Strength Cond. Res. 2021, 35, 411–419.
- Fritzen, A.M.; Lundsgaard, A.M.; Kiens, B. Tuning fatty acid oxidation in skeletal muscle with dietary fat and exercise. Nat. Rev. Endocrinol. 2020, 16, 683–696.
- Matsuoka, R.; Shirouchi, B.; Kawamura, S.; Baba, S.; Shiratake, S.; Nagata, K.; Imaizumi, K.; Sato, M. Dietary egg white protein inhibits lymphatic lipid transport in thoracic lymph duct-cannulated rats. J. Agric. Food Chem. 2014, 62, 10694–10700.
- Matsuoka, R. Health and nutritional function of egg protein. Amino Acid Res. 2019, 13, 27–32. (In Japanese)
- Matsuoka, R.; Kimura, M.; Arimitsu, K.; Kojima, T. New Health Functions of Egg White Protein (Lactic Fermented Egg White) and Its Application. J. Jpn. Soc. Nutr. Food Sci. 2021, 74, 147–154. (In Japanese)
- Sakai, K.; Ushiyama, Y.; Manabe, S. Peptic and pancreatic digestibility of raw and heat-treated hen’s egg white protein. Jpn. J. Pediatr. Allegy Clin. Immunol. 1999, 13, 36–42. (In Japanese)
- Handa, A.; Gennadios, A.; Hanna, M.A.; Weller, C.L.; Kuroda, N. Physical and molecular properties of egg-white lipid films. J. Food Sci. 1999, 64, 860–864.
- Takahashi, Y.; Matsuoka, R.; Kimura, M.; Masuda, Y.; Kunou, M.; Wada, Y.; Takahashi, Y.; Utsunomiya, K. Health function of egg white protein (No. 3: Mechanism for visceral fat lowering effects). In Proceedings of the 66th Annual Meeting of Japan Society of Nutrition and Food Science, Sendai, Japan, 18–20 May 2012; Komai, M., Ed.; Japan Society of Nutrition and Food Science: Tokyo, Japan, 2012.
- Rao, P.S.; Nolasco, E.; Handa, A.; Naldrett, M.J.; Alvarez, S.; Majumder, K. Effect of pH and heat treatment on the antioxidant activity of egg white protein-derived peptides after simulated in-vitro gastrointestinal digestion. Antioxidants 2020, 9, 1114.
- Yamada, Y.; Muraki, A.; Oie, M.; Kanegawa, N.; Oda, A.; Sawashi, Y.; Kaneko, K.; Yoshikawa, M.; Goto, T.; Takahashi, N.; et al. Soymorphin-5, a soy-derived mu-opioid peptide, decreases glucose and triglyceride levels through activating adiponectin and PPARalpha systems in diabetic KKAy mice. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E433–E440.
- Goto, T.; Mori, A.; Nagaoka, S. Soluble soy protein peptic hydrolysate stimulates adipocyte differentiation in 3T3-L1 cells. Mol. Nutr. Food Res. 2013, 57, 1435–1445.
- Kohno, M.; Hirotsuka, M.; Kito, M.; Matsuzawa, Y. Decreases in serum triacylglycerol and visceral fat mediated by dietary soybean beta-conglycinin. J. Atheroscler. Thromb. 2006, 13, 247–255.
- Ono, T.; Murakoshi, M.; Suzuki, N.; Iida, N.; Ohdera, M.; Iigo, M.; Yoshida, T.; Sugiyama, K.; Nishino, H. Potent anti-obesity effect of enteric-coated lactoferrin: Decrease in visceral fat accumulation in Japanese men and women with abdominal obesity after 8-week administration of enteric coated lactofferin tablets. Br. J. Nutr. 2010, 104, 1688–1695.
- Moriyama, T.; Kishimoto, K.; Nagai, K.; Urade, R.; Ogawa, T.; Utsumi, S.; Maruyama, N.; Maebashi, M. Soybean beta-conglycinin diet suppresses serum triglyceride levels in normal and genetically obese mice by induction of beta-oxidation, downregulation of fatty acid synthase, and inhibition of triglyceride absorption. Biosci. Biotechnol. Biochem. 2004, 68, 352–359.
- Matsuoka, R.; Kimura, M.; Muto, A.; Masuda, Y.; Sato, M.; Imaizumi, K. Mechanism for the cholesterol-lowering action of egg white protein in rats. Biosci. Biotechnol. Biochem. 2008, 72, 1506–1512.
- Mizuno, S.; Matsuura, K.; Gotou, T.; Nishimura, S.; Kajimoto, O.; Yabune, M.; Kajimoto, Y.; Yamamoto, N. Antihypertensive effect of casein hydrolysate in a placebo-controlled study in subjects with high-normal blood pressure and mild hypertension. Br. J. Nutr. 2005, 94, 84–91.
- Moro, T.; Brightwell, C.R.; Velarde, B.; Fry, C.S.; Nakayama, K.; Sanbongi, C.; Volpi, E.; Rasmussen, B.B. Whey Protein Hydrolysate Increases Amino Acid Uptake, mTORC1 Signaling, and Protein Synthesis in Skeletal Muscle of Healthy Young Men in a Randomized Crossover Trial. J. Nutr. 2019, 149, 1149–1158.
- Kwak, J.H.; Kim, M.; Lee, E.; Lee, S.H.; Ahn, C.W.; Lee, J.H. Effects of black soy peptide supplementation on blood pressure and oxidative stress: A randomized controlled trial. Hypertens. Res. 2013, 36, 1060–1066.
- Watabe, K. Function of egg white peptide “Peptifine®”. Jpn. Food Sci. 2013, 52, 17–24. (In Japanese)
- WHO. Energy and protein requirements (WHO technical report series, No. 724). In Report of a Joint FAO/WHO/UNU Expert Consultation; World Health Organization: Geneva, Switzerland, 1985.
- Seligson, F.H.; Rotruck, J.T. Tissue nonprotein sulfhydryl content and weight gain of rats as affected by dietary methionine level. J. Nutr. 1983, 13, 98–104.
- Steinberg, D.; Parthasarathy, S.; Carew, T.E.; Khoo, J.C.; Witztum, J.L. Beyond cholesterol. Modifications of low-density lipoprotein that increase its atherogenicity. N. Engl. J. Med. 1989, 320, 915–924.
- Matsumoto, K.; Koba, T.; Hamada, K.; Sakurai, M.; Higuchi, T.; Miyata, H. Branched-chain amino acid supplementation attenuates muscle soreness, muscle damage and inflammation during an intensive training program. J. Sports Med. Phys. Fitness 2009, 49, 424–431.
- Kato, Y.; Sawada, A.; Numao, S.; Miyauchi, R.; Imaizumi, K.; Sakamoto, S.; Suzuki, M. Effect of light resistance exercise after ingestion of a high-protein snack on plasma branched-chain amino acid concentrations in young adult females. J. Nutr. Sci. Vitaminol. 2009, 55, 106–111.
- Harada, R.; Taguchi, Y.; Urashima, K.; Sato, M.; Ohmori, T.; Morimatsu, F. Enhancement of Swimming Endurance in Mice by Chicken Breast Extract. Jpn. Soc. Nutr. Food. Sci. 2002, 55, 73–78. (In Japanese)
- Liu, J.; Wang, X.; Zhao, Z. Effect of whey protein hydrolysates with different molecular weight on fatigue induced by swimming exercise in mice. J. Sci. Food. Agr. 2014, 94, 126–130.
- Davalos, A.; Miguel, M.; Bartolome, B.; Lopez-Fandino, R. Antioxidant activity of peptides derived from egg white proteins by enzymatic hydrolysis. J. Food Prot. 2004, 67, 1939–1944.
- Sun, S.; Niu, H.; Yang, T.; Lin, Q.; Luo, F.; Ma, M. Antioxidant and anti-fatigue activities of egg white peptides prepared by pepsin digestion. J. Sci. Food Agr. 2014, 94, 3195–3200.
- Matsuoka, R.; Kimura, M.; Uno, S.; Shidara, H.; Kunou, M. Egg white hydrolysate improves fatigue due to short-term swimming load test in mice. Food Sci. Nutr. 2018, 6, 2314–2320.
- Sugiyama, K.; Kuriki, D.; Matsuoka, R.; Masuda, Y.; Kunou, M.; Daigo, E. Evaluation of the Functionality of Egg White Peptide as a Dietary Supplement for Sports. Nippon. Shokuhin Kagaku Kogaku Kaishi 2020, 67, 245–256. (In Japanese)
- Oe, M.; Sakamoto, H.; Nishiyama, H.; Sasahara, R.; Masuda, Y.; Adachi, M.; Nishiyama, T. Egg white hydrolyzate reduces mental fatigue: Randomized, double-blind, controlled study. BMC Res. Notes 2020, 13, 443.
This entry is offline, you can click here to edit this entry!