Developing varieties with diverse features that satisfy varied commercial needs, improving overall fruit quality, and quickly releasing them, are prerequisites in citrus breeding. However, these three goals require trade-offs in conventional breeding, even with the application of the marker-assisted selection technique. Conventional breeding cannot achieve these three goals simultaneously and it has been regarded as a breeding trilemma. Integrating a genomics-assisted breeding (GAB) approach that relies on quantitative trait locus detection by genome-wide association study and genome-wide prediction of a trait by genomic selection using enriched marker genotypes enhances breeding efficiency and contributes to eliminating the trilemma. Besides these efforts, the analysis of the genealogy of indigenous citrus varieties revealed that many high-quality indigenous varieties were selected within a few generations. It suggested that selecting a new premium quality hybrid is possible by integrating it with the GAB technique and helps avoid the trilemma. Researchers describes an ongoing comprehensive approach for integrating genomic-assisted breeding (GAB) with citrus genealogy on citrus breeding, called Citrus Breeding 2.0. This method can develop new cultivars with premium quality in a short period.
1. Introduction
Major citrus cultivars such as sweet orange, Clementine, lemon, grapefruit, and Satsuma have been produced worldwide for over 100 years, and are still considered economically important crops [
1,
2]. However, the demand has increased for high-quality new cultivars easy-to-peel and which meet consumers’ needs in a short period [
3]. Tremendous efforts have been devoted to mutation breeding in citrus to select a cultivar from sport, nucellar seedling [
4,
5,
6,
7], or after irradiation [
8]. Though mutation breeding has successfully selected spontaneous or induced mutants in various varieties based on color break, rind or flesh color, fruit shape, acidity, or sugar content, mutation occurs at a low frequency [
9], and the mutation spectrum depends on the varieties used for the method [
3,
5,
6,
7]. These limitations hamper the use of mutation breeding as a ubiquitous method for the development of new varieties. In contrast, crossbreeding is beneficial for developing novel, high-quality cultivars; however, in conventional breeding, it requires more time than mutation breeding, and the possibility to obtain a new elite hybrid depends on both the breeder’s experience and chance.
Crossbreeding is a systematic approach for developing new cultivars with novel traits using hybridization [
3,
10,
11]. However, it requires maintaining many seedlings in the orchard for an extended period and repeatedly evaluating their characteristics, and consequently this lengthy process is time-consuming and expensive. Therefore, in conventional breeding, satisfying fast breeding of a new hybrid, increasing fruit trait variation, and improving overall fruit quality are trade-offs, and it is difficult to achieve them simultaneously [
12]. This constraint is regarded as a ‘breeding trilemma’ that is difficult to avoid in conventional breeding [
13]. To solve these constraints in conventional breeding, the marker-assisted selection (MAS) technique has been introduced in citrus breeding [
12]. However, conventional MAS is limited to specific traits controlled by a few genes, and it is difficult to efficiently select various fruit traits that involve multiple genes in complex ways.
2. Citrus Crossbreeding Effort Aims to Improve Fruit Quality
2.1. Past Efforts of the Citrus Breeding Program in NARO
The fruit tree breeding program aims to develop new and unique high-quality cultivars. The citrus crossbreeding program of the Institute of Fruit Tree and Tea Science, NARO (NIFTS) started in 1946 and has produced hybrid cultivars intended for the domestic market [
14]. To date, the program has developed 39 hybrid cultivars with eight intermediary mother selections by repeatedly selecting a superior hybrid from a single cross. When the breeding program started, Satsuma, sweet orange, ponkan mandarin, and pummelo were typical in Japan, but each cultivar had drawbacks. For instance, although Satsuma is easy-to-peel, seedless, and productive, it has less flavor and a lower sugar content than oranges and exhibits alternate bearing. Sweet orange sets fruits with high sugar content and a strong and excellent aroma that is suitable for direct consumption and juice production. However, the rind of the sweet orange is hard to peel and requires a knife to eat. Ponkan mandarin has a preferred flavor and higher sugar content than Satsuma, but it has a puffy rind and contains many seeds that hamper eating. Pummelo is the generic name of a set of cultivars that produce large fruit with a thick yellow rind. These pummelo sets fruits with a high soluble solid content (SSC) and pleasant aroma. But they are difficult to peel, set many seeds, and the segment membrane is difficult to chew. The initial goal of the program was to develop a new cultivar with peelability comparable with Satsuma and a preferable aroma similar to sweet orange. ‘Kiyomi’ is a tangor selected from a cross between Satsuma and a sweet orange cross [
15]. ‘Kiyomi’ had better peelability than sweet orange, but not comparable to Satsuma, and while its aroma was superior to that of Satsuma, it was not comparable to sweet orange.
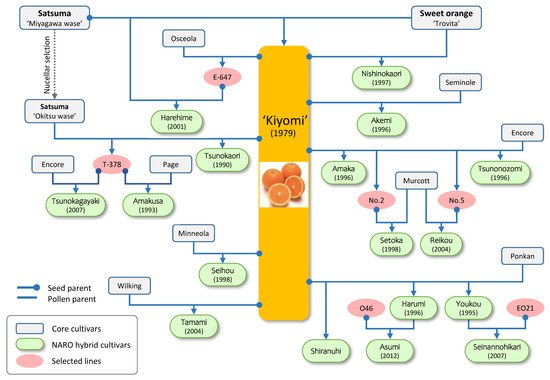
Besides its fruits, ‘Kiyomi’ has played a pivotal role in the breeding program, and it has been used as a mother plant because of its monoembryony (
Figure 1). ‘Kiyomi’ has contributed to the development of ten hybrid cultivars (‘Seihou’, ‘Tsunokaori’, ‘Youkou’, ‘Harumi’, ‘Akemi’, ‘Amaka’, ‘Nishinokaori’, ‘Tamami’, ‘Tsunonozomi’, and ‘Shiranuhi’), and six cultivars (‘Amakusa’, ‘Setoka’, ‘Harehime’, ‘Reikou’, ‘Tsunokagayaki’, and ‘Asumi’) are its progeny (
Figure 1 and
Table 1). These cultivars expanded the harvest season for five months (August to April) at open culture, enlarged fruit size (150–280 g), and improved SSC (>15). All of these cultivars except ‘Amakusa’, ‘Tamami’, and ‘Tsunonozomi’ set seedless fruit in open fields. Most of them are comparable to Satsuma in that they are easy to peel, and the segment membrane is easy to chew (
Table 1). Many of these cultivars have “good” flavor; among which, ‘Harehime’, ‘Tamami’ and ‘Asumi’ have a strong and excellent aroma similar to that of sweet orange. In addition, some of these cultivars have unique features for less rind puffing (‘Tsunokaori’, ‘Tsunonozomi’), less alternate bearing (‘Amakusa’), grenadine to red rind (‘Akemi’, ‘Reikou’), rich in beta-cryptoxanthin (‘Tsunokagayaki’, ‘Asumi’) and unique fruit shape (‘Shiranuhi’). All these cultivars are protected under the Plant Variety Protection and Seed Act, Japan except ‘Shiranuhi’ [
16], which is now produced in the USA (‘Sumo citrus’), Korea (‘Hallabong’), and Brazil (‘Kinsei’). Thus, starting from ‘Kiyomi’, a series of cultivars that have diverse and improved traits have been bred through several crossings. It demonstrates the performance of crossbreeding that enables the expansion of diversity and improvement of fruit quality.
Figure 1. Pedigree tree of ‘Kiyomi’ tangor with its offspring cultivars. Connector line terminating in a circle denotes seed parent, straight end denotes pollen parent.
This entry is adapted from the peer-reviewed paper 10.3390/horticulturae8060559