ANXA1 is found in the plasma membrane, cytoplasm, and nucleus, being described at low levels in the nuclear and cytoplasmic compartments of placental cells related to gestational diabetic diseases, and its translocation from the cytoplasm to the nucleus has been associated with a response to DNA damage. The approaches presented here open pathways for reflection upon, and intrinsic clarification of, the modulating action of this protein in the response to genetic material damage, as well as its level of expression and cellular localization.
- human placenta
- inflammation
- nuclear translocation
- gestational diabetes mellitus
- peptide Ac2-26
1. Introduction
2. ANXA1 and Inflammatory Processes
3. ANXA1 in the Placenta
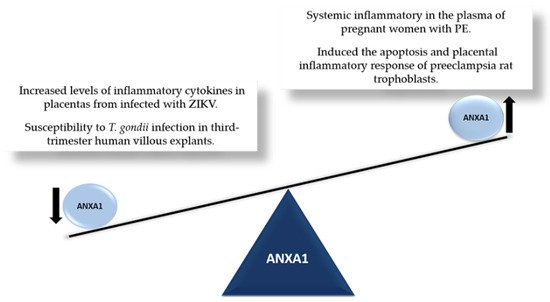
4. ANXA1 and Cell Survival
To survive and maintain genome integrity, organisms have DNA repair mechanisms that work effectively to remove lesions [68,69]. Quantitative proteomics studies indicate that ANXA1 may play a role in this DNA damage response [70]. Furthermore, in mammary adenocarcinoma cells (MCF7), it was shown that ANXA1 is related to protein cellular stress by protecting DNA against heat-induced damage [71].
Each type of DNA damage requires a specific set of cellular responses. Depending on the nature of the damage, different mechanisms are needed to repair the genetic material; when the damage exceeds the cell’s ability to repair itself, consequences such as the accumulation of mutations in the genome, or even cell death, may be observed [72,73]. Apoptosis and its associated regulatory mechanisms are crucial physiological events for the maintenance of placental homeostasis, and the imbalance of these processes can, among other consequences, compromise the function of the placenta, and therefore the success of the pregnancy [74].
Choi and collaborators developed an integrative network analysis to identify proteins that respond to the ATM inhibitor (a protein with a central role in the DNA damage signaling cascade) and physical interactions with DNA repair proteins. Interestingly, the analysis identified 53BP1 and ANXA1 as strong candidates. Complementing these results, the authors demonstrated that cell lines that do not express the ANXA1 protein are more sensitive to ionizing radiation [75].
Apoptosis is a physiological process of cell death in which cells undergo structural changes and are removed from the body without triggering an inflammatory response. The first indication of the involvement of ANXA1 in apoptosis was reported by McKanna, who showed that the expression of ANXA1 increased in alveolar cells of the mammary ducts undergoing apoptosis in post-lactational regression [76]. Subsequently, Sakamoto et al. reported that exogenous ANXA1 facilitated hydrogen peroxide-induced apoptosis in rat thymocytes [77]. Further evidence suggests that ANXA1 can mediate the proapoptotic effects of glucocorticoids in some cells, activating caspase-3 and acting on calcium fluxes [78,79].
5. ANXA1 in the Nucleus
ANXA1 has been found in the plasma membrane, cytoplasm, and nucleus [80,81]. Several studies have focused on the nuclear localization of ANXA1, and its translocation from the cytoplasm to the nucleus has been reported as a response to DNA damage, proliferative stimuli, and phosphorylation [82]. In addition, when overexpressed, intra- and/or extracellular ANXA1 translocated to the nucleus during apoptosis [83].
Studies indicate that ANXA1 nuclear translocation may be associated with cancer progression through the regulation of transcription factors and miRNAs [84], and the induction of apoptosis through the regulation of transcription factors such as p53 and p65 [85]. Recently, Luo et al. reported that ANXA1 determines the fate of retinal ganglion cells in a murine glaucoma model, and that its nuclear translocation induces apoptosis in these cells [86]. In addition, other works have shown that, when translocated to the nucleus, ANXA1 participates in neuronal apoptosis after cerebral ischemia [85,87]. In particular, the protein was found to act as a cofactor, binding to p53 in the nucleus and positively regulating its transcriptional activity, leading to the expression of the proapoptotic BID gene. Blocking ANXA1 nuclear translocation via a specific β-importin inhibitor reduced BID expression and inhibited the activation of the caspase-3 apoptotic pathway, attenuating neuronal apoptosis after ischemic stroke [87,88].
Although ANXA1 does not contain a classical nuclear localization signal, it has been observed that in the ANXA1 repeat domain III, amino acid residues from R228 to F237 function as a single nuclear translocation signal (NTS), and are required for the nuclear translocation of ANXA1 [82]. A recent study by Xia et al. found that the intracerebroventricular injection of the recombinant adenovirus vector S100A11 protects cells by preventing cerebral ischemia-induced neuronal cell apoptosis. Through the NTS, the adenovirus interacted directly with ANXA1, markedly decreasing its nuclear translocation [89].
Rhee and collaborators, aiming to identify whether the protein ANXA1 was related to cellular stress, observed that its gene expression levels increased in cells treated under stress conditions. Furthermore, in response, ANXA1 is translocated from the cytoplasm to the nucleus and perinuclear region. Its role in resolving stress-induced transcriptional activation was investigated, and the associated alteration was significantly larger than in cells maintained under different conditions [90].
The presence of ANXA1 in the nucleus has also been suggested as a significant predictor of survival in oral and esophageal squamous cell carcinomas. It was observed that ANXA1 expression, although decreased in the cytosol and membranes, was increased in the nuclei of esophageal cancer cells. Furthermore, patients with low nuclear ANXA1 expression had better prognoses than those with high protein expression [67,91]. Similar studies showed that ANXA1 is expressed in both gastric adenocarcinoma and normal tissues. In gastric adenocarcinoma tissues, ANXA1 is expressed in both the cytoplasm and the nucleus, and its nuclear location correlates with the advanced stage of the disease and peritoneal dissemination [92]. Moreover, it was demonstrated in L5178Y tk+/− mouse lymphoma cells treated with DNA-damaging agents that the quantity of nuclear ANXA1 increased while cytoplasmic ANXA1 levels decreased, suggesting that nuclear translocation of this protein occurs in response to the signaling of damaged DNA [93].
Considered in combination, these findings from the literature indicate that, in addition to its level of expression, the subcellular localization and translocation of ANXA1 may play an important role in several pathologies.
6. Perspective and Conclusions
Since its discovery as an anti-phospholipase protein, ANXA1 has been found to exhibit a wide range of anti-inflammatory properties. However, further research is needed to define the processes and factors that influence its nuclear translocation, with the aim of identifying the mechanisms by which this protein performs its functions in the nucleus.
Although this discussion has focused on the placenta, this review provides novel insights into how ANXA1 regulates the body’s pathophysiological processes, predominantly in relation to its nuclear action. This aspect will be particularly important for further investigation into the role of ANXA1 in the nucleus, and the development of new inflammatory therapies based on the understanding and targeting of this protein.
This entry is adapted from the peer-reviewed paper 10.3390/cells11132057