Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Vitamin B6 is a cofactor for approximately 150 reactions that regulate the metabolism of glucose, lipids, amino acids, DNA, and neurotransmitters. In addition, it plays the role of antioxidant by counteracting the formation of reactive oxygen species (ROS) and advanced glycation end-products (AGEs). Epidemiological and experimental studies indicated an evident inverse association between vitamin B6 levels and diabetes, as well as a clear protective effect of vitamin B6 on diabetic complications.
- vitamin B6
- diabetes
- AGEs
1. Vitamin B6: Roles and Synthesis
Vitamin B6 is a very important compound for general cellular metabolism [1]. The term vitamin B6 refers to six common forms or vitamers, namely pyridoxine (PN), pyridoxal (PL), pyridoxamine (PM), and their related 5′-phosphate derivatives (PNP, PLP, and PMP). The biologically active form, the pyridoxal 5′-phosphate (PLP), acts as coenzyme in about 150 distinct enzymatic activities that catalyze crucial metabolic reactions, such as synthesis, transformation, and degradation of amines and amino acids, supply of one carbon units, transsulfuration, synthesis of tetrapyrrolic compounds (including heme) and polyamines, biosynthesis, and degradation of neurotransmitters [2][3]. Although vitamin B6 is not classified as a classical antioxidant compound, it is able to quench oxygen reactive species (ROS) [4] and counteract the formation of advanced glycation end products (AGEs), genotoxic compounds associated with senescence, and diabetes [5]. Furthermore, PLP works as a modulator of transcription factors, has a role in enzyme folding, and can bind to steroid receptors, playing a role in membrane transport [6].
Mammals, different from microorganisms, are not able to synthesize PLP but recycle it through a salvage pathway from B6 vitamers as pyridoxal (PL), pyridoxamine (PM), and pyridoxine (PN) contained in food [7]. In the cytoplasm PL, PM, and PN are converted into the 5′-phosphorylated vitamers through pyridoxal kinase (PDXK), while the FMN-dependent pyridoxine 5′-phosphate oxidase (PNPO) converts PNP and PMP into PLP (Figure 1). Once ingested, PLP, PNP, and PMP are dephosphorylated by the tissue-non-specific alkaline phosphatase (TNSALP), which is anchored to the cell membrane. Then, PM, PN, and PL are absorbed from the upper small intestine by a carrier-mediated system and is delivered to the liver. Here, they are converted to PLP, thanks to the combined action of PDXK and PNPO. From the liver, PLP bound to albumin, along with dephosphorylated B6 vitamers, reach the peripheral tissues through the blood stream. In order to enter cells, PLP needs to be dephosphorylated again by TNSALP [8]. In the cytosol, a ubiquitous PLP phosphatase is instead required for vitamin B6 catabolism [9].
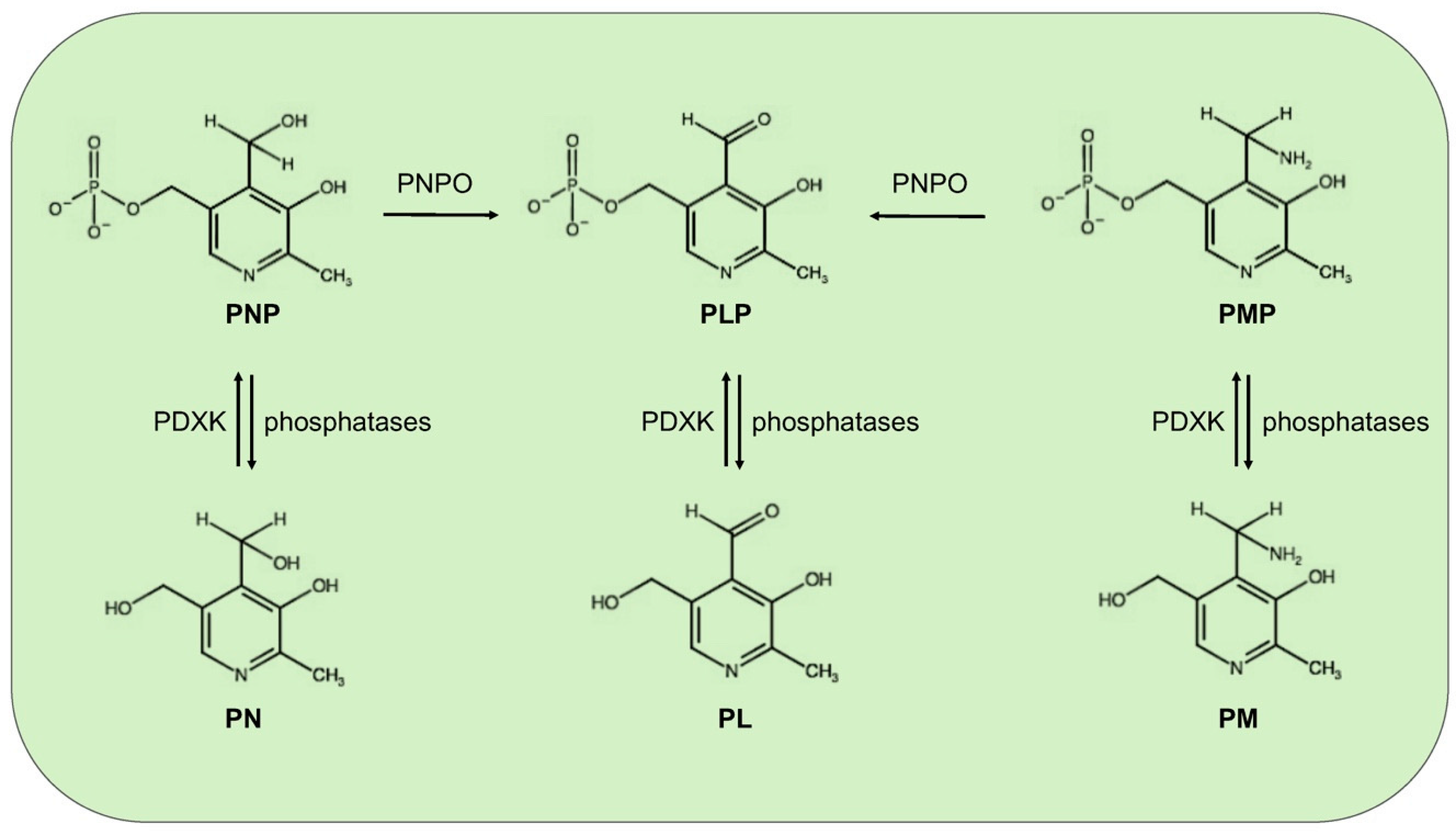
Figure 1. Schematic vitamin B6 metabolism in humans. The diagram corresponds to the pyridoxal 5′-phosphate salvage pathway. PLP, pyridoxal 5′-phosphate; PNP, pyridoxine 5′-phosphate; PMP, pyridoxamine 5′-phosphate; PL, pyridoxal; PN, pyridoxine; PM, pyridoxamine; PDXK, pyridoxal kinase; and PNPO, pyridoxine 5′-phosphate oxidase.
Vitamin B6 recommended dietary allowance is 1.3 mg day-1 for adults; it is present in several foods including meat, fish, poultry, vegetables, and fruits, thus, a severe deficiency of this vitamin is uncommon in developed countries, within any diet. However, PLP concentrations appear to be reduced in certain contexts, as for example, alcoholism [10], obesity [11], and pregnancy [12]. Moreover, some pathological conditions such as end-stage renal diseases, chronic renal insufficiency, and other kidney diseases can lower the vitamin B6 levels [12]. In addition, vitamin B6 deficiency has been correlated to malabsorption syndromes, such as celiac disease and inflammatory bowel diseases [12][13][14]. Even the intake of certain medications, including isoniazid [15], penicillamine [16], and cycloserine [17], as well as oral contraceptives, can reduce PLP availability [18]. PLP levels can also be decreased by the inflammation conditions and stress hormones [19][20].
2. Vitamin B6 and Diabetes
By considering the plethora of reactions in which vitamin B6 is involved, it is not surprising that its deficiency has been implicated in several clinically relevant diseases, including autism, schizophrenia, Alzheimer, Parkinson, epilepsy, Down’s syndrome, diabetes, and cancer; however, the underlying mechanisms remain unknown in most cases [21][22][23].
Diabetes mellitus (DM) represents a global health problem, touching more than 400 million people and consists of a group of metabolic disorders characterized by persistent hyperglycemia arising from impaired insulin secretion, insulin action, or both [24]. DM is a multifactorial disease caused by the concerted action of genetic and environmental factors, and on the basis of its etiology, it can be classified into three major types—type1 (T1D), type2 (T2D), and gestational diabetes (GDM). T1D is an autoimmune disorder that leads to the destruction of pancreatic beta-cells and accounts for only 5–10% of all diabetes. T2D, the more frequent form (90–95%), is mainly caused by insulin resistance consisting of a diminished tissue response to insulin that leads glucose to accumulate in blood. Consequently, the rate of insulin secretion increases to meet the body’s needs, but this overload, in the long-term, compromises pancreas functionality. GDM is a common pregnancy complication that affects approximately 14% of pregnancies worldwide. It is associated with insulin resistance, in turn generated by a combined action of pregnancy hormones and other factors [25].
Substantial evidence correlates vitamin B6 to diabetes and its complications. Some population screenings have been carried out to compare PLP levels in diabetic groups vs. healthy subjects; in addition, several studies focused on the impact of vitamin B6 on diabetic complications and others on the effectiveness of vitamin B6 as a preventive treatment. Vitamin B6 levels are commonly assessed by measuring plasma pyridoxal 5′-phosphate (PLP) concentration and an inadequate vitamin B6 status is generally associated with a concentration, under the cut-off level of 30 nmol/L. Other methods include the measurement of plasma pyridoxal or total vitamin B6 and urinary 4-pyridoxic acid, as well the ratio between PLP and PL [26]. By examining the studies reported in literature, an inverse relation between vitamin B6 levels and diabetes emerges. Satyanarayana and coworkers [27] in a cross-sectional case-control study found that the mean plasma PLP levels were significantly lower in T2D subjects, compared to the healthy controls. By comparing the results obtained in a Korean study by Ahn and coworkers [28] to those obtained by Nix and collaborators [29] in a German cohort, vitamin B6 levels appeared to be inversely related to the progression of diabetes. Ahn and collaborators [28], in fact, examined diabetic people with an early status of the disease, finding a mean plasma PLP level reduction to be relevant but not statistically significant, with respect to controls; in contrasts, the diabetic group examined by Nix [29], being composed of people with advanced clinical stage, exhibited median plasma concentrations of PLP, PN, and PL that were significantly decreased in a diabetic group compared to the controls. Interestingly, median plasma levels of the PM, PMP, and pyridoxic acid were significantly higher in the diabetes groups than in the controls; this finding led Nix and collaborators to advance the hypothesis that T2D might be associated with an altered activity of the enzymes involved in the interconversion of B6 vitamers [29]. In another study, based on the evidence of increased urinary clearance of vitamin B6, it was hypothesized that decreased vitamin B6 levels in T2D subjects could derive from an impaired reabsorption processes [30]. The same inverse relationship between B6 levels and diabetes was observed in experimental animals [31][32]. Roger was the first to describe decreased PLP levels in streptozotocin-diabetic rats accompanied by less storage in the liver of the mitochondrial PLP [31].
Decreased PLP levels have also be associated with GDM. In a study performed in a group of women affected by GDM, Bennink and Schreurs [33] found that 13 out of 14 displayed reduced PLP levels. Moreover, pyridoxine administration ameliorated oral glucose tolerance. Analogous results were obtained by Spellacy and coworkers [34], which found a clear blood glucose decrease and a normalization of insulin secretion following pyridoxine therapy in GDM women, indicating that vitamin B6 might ameliorate plasma insulin biological activity.
Other intervention studies reported that pyridoxine supplementation is capable of lowering blood glucose levels in streptozotocin-treated rats [35], as well as glycosylated hemoglobin levels in T2D patients [36]. Moreover, Kim and collaborators [37] showed that vitamin B6 can reduce postprandial blood glucose levels following sucrose and starch ingestion, by inhibiting the activity of small-intestinal α-glucosidases.
3. Vitamin B6 and DNA Damage in Diabetes
It is widely accepted that oxidative stress is a major risk factor for onset and progression of diabetes. Many of the common risk factors, including obesity and unhealthy eating habits, contribute to produce an oxidative environment that might increase insulin resistance or impair glucose tolerance. Hyperglycemia, in turn, contributes to progression and maintenance of an overall oxidative environment [38]. Consequently, diabetes was associated with reduced levels of antioxidants, such as GSH, vitamin C, and vitamin E [39], and to a low efficiency of DNA repair systems [40][41].
Evidence indicated that both T1D and T2D diabetic patients exhibit oxidative damage and DNA strand breaks [42][43]. It was shown that tissues from diabetic rats and the urine of T1D and T2D patients have increased levels of 8-Oxo-7,8-dihydro-2′-deoxyguanosine (8-oxodG), a sensitive marker of reactive oxygen species (ROS)-induced DNA damage [44][45][46]. In addition, diabetic patients have a higher frequency of sister chromatid exchange than healthy subjects [47][48]. Another study reported high levels of stable chromosomal aberrations in peripheral lymphocytes associated with T2D, and directly correlated with the risk of early diabetes-related death [48]. Moreover, an elevated frequency of micronuclei was found in T2D patients with no microvascular or macrovascular complications [49]. Recently, micronuclei frequency was positively related to glycated hemoglobin (HbA1c) levels, as well as to fasting plasma glucose in T2D [50].
By considering that PLP is both an antioxidant molecule and a cofactor for enzymes involved in DNA metabolism, it is expected that its reduced availability can contribute to increase DNA damage associated with diabetes. Studies in Drosophila and human cell cultures helped to gain evidence in support of this hypothesis, demonstrating that PLP deficiency can cause DNA damage throughout the formation of AGEs [51][52]. PLP deficiency in Drosophila and human cultured cells, trigger the formation of chromosome aberrations (CABs). In particular, mutations in the pyridoxal kinase gene (dPdxk1) as well as the RNAi-induced silencing of pyridoxine 5′-phosphate oxidase gene (sgll), produced CABs in brain cells [51][52]. Similarly, CABs were produced in wild-type individuals through PLP inhibitors, such as 4-DP, cycloserine, isoniazid, and penicillamine [51]. Interestingly, in all of these cases, CAB frequency was strongly enhanced by treatment with sugars, such as glucose, fructose, or sucrose, whereas in contrast, sugar treatments did not induce CABs in wild-type brains. In addition to providing the first evidence that correlates vitamin B6, DNA damage, and hyperglycemia, these data [51][52] also indicate that low PLP levels and high glucose might synergize in the process of CAB formation. Notably, PLP-depleted cells accumulated significant levels of AGEs, whose formation is enhanced by glucose treatment [51][52]. Remarkably, the treatment with the antioxidant alpha-lipoic acid, rescued not only AGEs but also CABs [51][52], further highlighting the cause–effect relationship between high glucose and CABs in low PLP conditions. These data allowed building a model, according to which decreased PLP levels induce hyperglycemia, which in turn produces AGEs that are responsible for DNA damage through ROS formation. This model could also be applied to humans, based on the following considerations—(1) the human PDXK gene inserted in dPdxk1 mutant flies is capable of reducing hyperglycemia, CABs, and AGE accumulation [51]; (2) RNAi-induced silencing of the human PDXK gene produces CABs in fibroblasts and HeLa cells, as well as 4-DP treatment in mock cells [51]; (3) alpha-lipoic acid treatment rescued DNA damage induced by PDXK depletion [51]; (4) the expression in the dPdxk1 mutant flies of 4 PDXK human variants with reduced or impaired catalytic activity did not rescue CABs nor AGEs [53]. Taken together, these data suggest not only that low PLP levels could contribute to produce DNA damage in diabetic cells throughout AGE formation, but more importantly, that diabetic patients need to monitor their PLP content to avoid DNA damage, which is a well-known cancer prerequisite.
4. Vitamin B6 Diabetes and Cancer
Diabetic patients have an increased risk for developing different types of tumors, including liver, pancreas, colorectal, and lung cancers, but the underlying mechanisms are not fully elucidated [54][55][56]. Hyperinsulinemia and hyperglycemia were hypothesized to have a role in mediating this association, mainly by promoting cell growth [57]. However, it is thought that hyperglycemia might also impact cancer by triggering DNA damage through AGEs and ROS formation.
Based on the above-mentioned findings obtained in Drosophila, by considering that DNA damage caused by PLP deficiency is sugar-sensitive, it is reasonable to suppose that in a given diabetic context, the combined action of reduced PLP availability and high endogenous glucose levels could increase the severity of DNA damage. Evidence supporting this hypothesis was provided by studies in Drosophila. It was demonstrated in fact that treatment with 4-DP resulted in much more severe DNA damage in diabetic individuals than in wild-type flies [58]. In particular, brains from two different fly models of T2D displayed 60–80% of CABs (vs. 25% in 4-DP treated wild-type individuals) and accumulated many more AGEs. Additionally, treatment with alpha-lipoic acid rescued both AGEs and CABs, confirming that CABs were largely produced by AGEs [58]. Consequently, extrapolated to humans, this finding indicates that low PLP levels might contribute to increased cancer-risk in diabetic patients. It is possible, in fact, to envisage that in an oxidative environment in which antioxidant defenses and DNA repair are weakened, a decline in the capability to counteract AGEs and ROS induced by low PLP levels, might cause an amplification of genotoxic effects, increasing the risk for developing cancer.
This entry is adapted from the peer-reviewed paper 10.3390/ijms21103669
References
- Hellmann, H.; Mooney, S. Vitamin B6: A molecule for human health? Molecules 2010, 15, 442–459.
- Percudani, R.; Peracchi, A. A genomic overview of pyridoxal-phosphate-dependent enzymes. EMBO Rep. 2003, 4, 850–854.
- Di Salvo, M.L.; Contestabile, R.; Safo, M.K. Vitamin B(6) salvage enzymes: Mechanism, structure and regulation. Biochim. Biophys. Acta 2011, 1814, 1597–1608.
- Bilski, P.; Li, M.Y.; Ehrenshaft, M.; Daub, M.E.; Chignell, C.F. Vitamin B6 (pyridoxine) and its derivatives are efficient singlet oxygen quenchers and potential fungal antioxidants. Photochem. Photobiol. 2000, 71, 129–134.
- Booth, A.A.; Khalifah, R.G.; Todd, P.; Hudson, B.G. In vitro kinetic studies of formation of antigenic advanced glycation end products (AGEs). Novel inhibition of post-Amadori glycation pathways. J. Biol. Chem. 1997, 272, 5430–5437.
- Lambrecht, G.; Braun, K.; Damer, M.; Ganso, M.; Hildebrandt, C.; Ullmann, H.; Kassack, M.U.; Nickel, P. Structure-activity relationships of suramin and pyridoxal-5′-phosphate derivatives as P2 receptor antagonists. Curr. Pharm. Des. 2002, 8, 2371–2399.
- McCormick, D.B. Two interconnected B vitamins: Riboflavin and pyridoxine. Physiol. Rev. 1989, 69, 1170–1198.
- Said, H.M. Recent advances in carrier-mediated intestinal absorption of water-soluble vitamins. Annu. Rev. Physiol. 2004, 66, 419–446.
- Jang, Y.M.; Kim, D.W.; Kang, T.C.; Won, M.H.; Baek, N.I.; Moon, B.J.; Choi, S.Y.; Kwon, O.S. Human pyridoxal phosphatase. Molecular cloning, functional expression, and tissue distribution. J. Biol. Chem. 2003, 278, 50040–50046.
- Cravo, M.L.; Camilo, M.E. Hyperhomocysteinemia in chronic alcoholism: Relations to folic acid and vitamins B(6) and B(12) status. Nutrition 2000, 16, 296–302.
- Ferro, Y.; Carè, I.; Mazza, E.; Provenzano, F.; Colica, C.; Torti, C.; Romeo, S.; Pujia, A.; Montalcini, T. Protein and vitamin B6 intake are associated with liver steatosis assessed by transient elastography, especially in obese individuals. Clin. Mol. Hepatol. 2017, 23, 249–259.
- Merrill, A.H., Jr.; Henderson, J.M. Diseases associated with defects in vitamin B6 metabolism or utilization. Annu. Rev. Nutr. 1987, 7, 137–156.
- Kowlessar, O.D.; Haeffner, L.J.; Benson, G.D. Abnormal tryptophan metabolism in patients with adult celiac disease, with evidence for deficiency of vitamin B6. J. Clin. Investig. 1964, 43, 894–903.
- Chiang, E.P.; Selhub, J.; Bagley, P.J.; Dallal, G.; Roubenoff, R. Pyridoxine supplementation corrects vitamin B6 deficiency but does not improve inflammation in patients with rheumatoid arthritis. Arthritis Res. Ther. 2005, 7, R1404-11.
- Biehl, J.P.; Vilter, R.W. Effect of isoniazid on vitamin B6 metabolism; its possible significance in producing isoniazid neuritis. Proc. Soc. Exp. Biol. Med. 1954, 85, 389–392.
- Jaffe, I.A.; Altman, K.; Merryman, P. The antipyridoxine effect of penicillamine in man. J. Clin. Investig. 1964, 43, 1869–1873.
- Yamada, K.; Sawaki, S.; Hayami, S. Inhibitory effect of cycloserine on some enzymic activities related to vitamin B6. J. Vitaminol. 1957, 3, 68–72.
- Lussana, F.; Zighetti, M.L.; Bucciarelli, P.; Cugno, M.; Cattaneo, M. Blood levels of homocysteine, folate, vitamin B6 and B12 in women using oral contraceptives compared to non-users. Thromb. Res. 2003, 112, 37–41.
- Oxenkrug, G.F. Tryptophan kynurenine metabolism as a common mediator of genetic and environmental impacts in major depressive disorder: The serotonin hypothesis revisited 40 years later. Isr. J. Psychiatry Relat. Sci. 2010, 47, 56–63.
- Midttun, O.; Ulvik, A.; Ringdal Pedersen, E.; Ebbing, M.; Bleie, O.; Schartum-Hansen, H.; Nilsen, R.M.; Nygård, O.; Ueland, P.M. Low plasma vitamin B-6 status affects metabolism through the kynurenine pathway in cardiovascular patients with systemic inflammation. J. Nutr. 2011, 141, 611–617.
- Di Salvo, M.L.; Safo, M.K.; Contestabile, R. Biomedical aspects of pyridoxal 5′-phosphate availability. Front. Biosci. 2012, 4, 897–913.
- Merigliano, C.; Mascolo, E.; Burla, R.; Saggio, I.; Vernì, F. The Relationship Between Vitamin B6, Diabetes and Cancer. Front. Genet. 2018, 9, 388.
- Contestabile, R.; di Salvo, M.L.; Bunik, V.; Tramonti, A.; Vernì, F. The multifaceted role of vitamin B(6) in cancer: Drosophila as a model system to investigate DNA damage. Open Biol. 2020, 10, 200034.
- American Diabetes Association. Diagnosis and classification of diabetes mellitus. Diabetes Care 2013, 36 (Suppl. 1), S67–S74.
- Plows, J.F.; Stanley, J.L.; Baker, P.N.; Reynolds, C.M.; Vickers, M.H. The Pathophysiology of Gestational Diabetes Mellitus. Int. J. Mol. Sci. 2018, 19, 3342.
- Leklem, J.E. Vitamin B-6: A status report. J. Nutr. 1990, 120 (Suppl. 11), 1503–1507.
- Satyanarayana, A.; Balakrishna, N.; Pitla, S.; Reddy, P.Y.; Mudili, S.; Lopamudra, P.; Suryanarayana, P.; Viswanath, K.; Ayyagari, R.; Reddy, G.B. Status of B-vitamins and homocysteine in diabetic retinopathy: Association with vitamin-B12 deficiency and hyperhomocysteinemia. PLoS ONE 2011, 6, e26747.
- Ahn, H.J.; Min, K.W.; Cho, Y.O. Assessment of vitamin B(6) status in Korean patients with newly diagnosed type 2 diabetes. Nutr. Res. Pract. 2011, 5, 34–39.
- Nix, W.A.; Zirwes, R.; Bangert, V.; Kaiser, R.P.; Schilling, M.; Hostalek, U.; Obeid, R. Vitamin B status in patients with type 2 diabetes mellitus with and without incipient nephropathy. Diabetes Res. Clin. Pract. 2015, 107, 157–165.
- Iwakawa, H.; Nakamura, Y.; Fukui, T.; Fukuwatari, T.; Ugi, S.; Maegawa, H.; Doi, Y.; Shibata, K. Concentrations of Water-Soluble Vitamins in Blood and Urinary Excretion in Patients with Diabetes Mellitus. Nutr. Metab. Insights 2016, 9, 85–92.
- Rogers, K.S.; Higgins, E.S.; Kline, E.S. Experimental diabetes causes mitochondrial loss and cytoplasmic enrichment of pyridoxal phosphate and aspartate aminotransferase activity. Biochem. Med. Metab. Biol. 1986, 36, 91–97.
- Okada, M.; Shibuya, M.; Yamamoto, E.; Murakami, Y. Effect of diabetes on vitamin B6 requirement in experimental animals. Diabetes Obes. Metab. 1999, 1, 221–225.
- Bennink, H.J.; Schreurs, W.H. Improvement of oral glucose tolerance in gestational diabetes by pyridoxine. Br. Med. J. 1975, 3, 13–15.
- Spellacy, W.N.; Buhi, W.C.; Birk, S.A. Vitamin B6 treatment of gestational diabetes mellitus: Studies of blood glucose and plasma insulin. Am. J. Obstet. Gynecol. 1977, 127, 599–602.
- Nair, A.R.; Biju, M.P.; Paulose, C.S. Effect of pyridoxine and insulin administration on brain glutamate dehydrogenase activity and blood glucose control in streptozotocin-induced diabetic rats. Biochim. Biophys. Acta 1998, 1381, 351–354.
- Solomon, L.R.; Cohen, K. Erythrocyte O2 transport and metabolism and effects of vitamin B6 therapy in type II diabetes mellitus. Diabetes 1989, 38, 881–886.
- Kim, H.H.; Kang, Y.R.; Lee, J.Y.; Chang, H.B.; Lee, K.W.; Apostolidis, E.; Kwon, Y.I. The Postprandial Anti-Hyperglycemic Effect of Pyridoxine and Its Derivatives Using In Vitro and In Vivo Animal Models. Nutrients 2018, 10, 285.
- Rains, J.L.; Jain, S.K. Oxidative stress, insulin signaling, and diabetes. Free Radic. Biol. Med. 2011, 50, 567–575.
- Bravi, M.C.; Armiento, A.; Laurenti, O.; Cassone-Faldetta, M.; De Luca, O.; Moretti, A.; De Mattia, G. Insulin decreases intracellular oxidative stress in patients with type 2 diabetes mellitus. Metabolism 2006, 55, 691–695.
- Blasiak, J.; Arabski, M.; Krupa, R.; Wozniak, K.; Zadrozny, M.; Kasznicki, J.; Zurawska, M.; Drzewoski, J. DNA damage and repair in type 2 diabetes mellitus. Mutat. Res. 2004, 554, 297–304.
- Zhong, A.; Chang, M.; Yu, T.; Gau, R.; Riley, D.J.; Chen, Y.; Chen, P.L. Aberrant DNA damage response and DNA repair pathway in high glucose conditions. J. Cancer Res. Updates 2018, 7, 64–74.
- Goodarzi, M.T.; Navidi, A.A.; Rezaei, M.; Babahmadi-Rezaei, H. Oxidative damage to DNA and lipids: Correlation with protein glycation in patients with type 1 diabetes. J. Clin. Lab. Anal. 2010, 24, 72–76.
- Tatsch, E.; Bochi, G.V.; Piva, S.J.; De Carvalho, J.A.; Kober, H.; Torbitz, V.D.; Duarte, T.; Signor, C.; Coelho, A.C.; Duarte, M.M.; et al. Association between DNA strand breakage and oxidative, inflammatory and endothelial biomarkers in type 2 diabetes. Mutat. Res. 2012, 732, 16–20.
- Umemura, T.; Sai, K.; Takagi, A.; Hasegawa, R.; Kurokawa, Y. Formation of 8-hydroxydeoxyguanosine (8-OH-dG) in rat kidney DNA after intraperitoneal administration of ferric nitrilotriacetate (Fe-NTA). Carcinogenesis 1990, 11, 345–347.
- Dandona, P.; Thusu, K.; Cook, S.; Snyder, B.; Makowski, J.; Armstrong, D.; Nicotera, T. Oxidative damage to DNA in diabetes mellitus. Lancet 1996, 347, 444–445.
- Hinokio, Y.; Suzuki, S.; Hirai, M.; Chiba, M.; Hirai, A.; Toyota, T. Oxidative DNA damage in diabetes mellitus: Its association with diabetic complications. Diabetologia 1999, 42, 995–998.
- Binici, D.N.; Karaman, A.; Coşkun, M.; Oğlu, A.U.; Uçar, F. Genomic damage in patients with type-2 diabetes mellitus. Genet. Couns. 2013, 24, 149–156.
- Boehm, B.O.; Möller, P.; Högel, J.; Winkelmann, B.R.; Renner, W.; Rosinger, S.; Seelhorst, U.; Wellnitz, B.; März, W.; Melzner, J.; et al. Lymphocytes of type 2 diabetic women carry a high load of stable chromosomal aberrations: A novel risk factor for disease-related early death. Diabetes 2008, 57, 2950–2957.
- Martínez-Pérez, L.M.; Cerda-Flores, R.M.; Gallegos-Cabriales, E.C.; Dávila-Rodríguez, M.I.; Ibarra-Costilla, E.; Cortés-Gutiérrez, E.I. Frequency of micronuclei in Mexicans with type 2 diabetes mellitus. Prague Med. Rep. 2007, 108, 248–255.
- Grindel, A.; Brath, H.; Nersesyan, A.; Knasmueller, S.; Wagner, K.H. Association of Genomic Instability with HbA1c levels and Medication in Diabetic Patients. Sci. Rep. 2017, 7, 41985.
- Marzio, A.; Merigliano, C.; Gatti, M.; Vernì, F. Sugar and chromosome stability: Clastogenic effects of sugars in vitamin B6-deficient cells. PLoS Genet. 2014, 10, e1004199.
- Mascolo, E.; Amoroso, N.; Saggio, I.; Merigliano, C.; Vernì, F. Pyridoxine/pyridoxamine 5′-phosphate oxidase (Sgll/PNPO) is important for DNA integrity and glucose homeostasis maintenance in Drosophila. J. Cell. Physiol. 2020, 235, 504–512.
- Mascolo, E.; Barile, A.; Mecarelli, L.S.; Amoroso, N.; Merigliano, C.; Massimi, A.; Saggio, I.; Hansen, T.; Tramonti, A.; Di Salvo, M.L.; et al. The expression of four pyridoxal kinase (PDXK) human variants in Drosophila impacts on genome integrity. Sci. Rep. 2019, 9, 14188.
- Mocellin, S.; Briarava, M.; Pilati, P. Vitamin B6 and Cancer Risk: A Field Synopsis and Meta-Analysis. J. Natl. Cancer Inst. 2017, 109, 1–9.
- Zuo, H.; Ueland, P.M.; Midttun, Ø.; Tell, G.S.; Fanidi, A.; Zheng, W.; Shu, X.; Xiang, Y.; Wu, J.; Prentice, R.; et al. Vitamin B6 catabolism and lung cancer risk: Results from the Lung Cancer Cohort Consortium (LC3). Ann. Oncol. 2019, 30, 478–485.
- Gylling, B.; Myte, R.; Schneede, J.; Hallmans, G.; Häggström, J.; Johansson, I.; Ulvik, A.; Ueland, P.M.; Van Guelpen, B.; Palmqvist, R. Vitamin B-6 and colorectal cancer risk: A prospective population-based study using 3 distinct plasma markers of vitamin B-6 status. Am. J. Clin. Nutr. 2017, 105, 897–904.
- Vigneri, P.; Frasca, F.; Sciacca, L.; Pandini, G.; Vigneri, R. Diabetes and cancer. Endocr. Relat. Cancer 2009, 16, 1103–1123.
- Merigliano, C.; Mascolo, E.; La Torre, M.; Saggio, I.; Vernì, F. Protective role of vitamin B6 (PLP) against DNA damage in Drosophila models of type 2 diabetes. Sci. Rep. 2018, 8, 11432.
This entry is offline, you can click here to edit this entry!