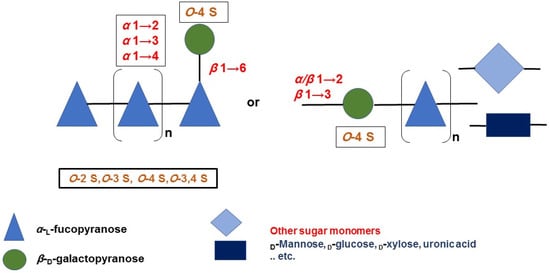
Fucoidans encompass versatile and heterogeneous sulfated biopolysaccharides of marine origin, specifically brown algae and marine invertebrates. The reported studies revealed diverse chemical skeletons in which l-fucose is the main sugar monomer. However, other sugars, i.e., galactose, mannose, etc., have been identified to be interspersed, forming several heteropolymers, including galactofucans/fucogalactans (G-fucoidans). Particularly, sulfated galactofucans are associated with rich chemistry contributing to more promising bioactivities than fucans and other marine polysaccharides. The previous reports in the last 20 years showed that G-fucoidans derived from Undaria pinnatifida were the most studied; 21 bioactivities were investigated, especially antitumor and antiviral activities, and unique biomedical applications compared to other marine polysaccharides were demonstrated.
- bioactives
- brown seaweeds
- fucoidans
- heteropolysaccharides
- structural features
- sulfated galactofucans
1. Introduction
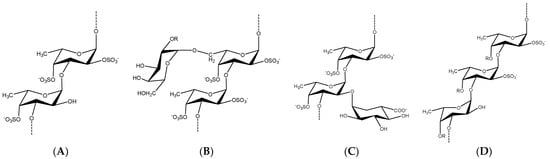
2. Occurrence, Distribution, and Chemistry
|
Brown Algae (Seaweed) Species |
Source of Seaweed Biomass |
Structural Characteristics |
References |
|||||
|---|---|---|---|---|---|---|---|---|
|
Monosaccharide Composition |
Glycosidic Bonds of Backbone |
Molecular Weight (kDa) |
Fucose/Galactose Ratio |
Sulfate Content (%) |
Sulfation Pattern |
|||
|
Dictyotales |
||||||||
|
Canistrocarpus cervicornis |
Wild |
Gal, fuc, glcAc, xyl, |
ND |
2 |
16.5 |
ND |
[67] |
|
|
Dictyota dichotoma |
Wild |
Gal, fuc, man, xyl, ara, rha, glc |
23.6 |
1.5 |
33 |
ND |
[68] |
|
|
D. implexa |
Wild |
Gal, fuc |
ND |
1 |
18.3 |
ND |
[69] |
|
|
Lobophora variegata |
Wild |
Gal, fuc, Glc, man, xyl, glcAc; Gal, fuc; Gal, fuc, Glc |
(1,3)- and (1,4)-α-l-fuc, and (1,3)-β-d-gal |
35; ND; 1400 |
0.79; 0.5; 0.5 |
32.6; 0.2 *;15 |
At C4 (fuc) |
|
|
L. variegata |
ND |
Gal, fuc |
ND |
360–1600 |
0.3 |
23.3–35.5 |
ND |
[71] |
|
Padina boryana |
Wild |
Gal, fuc |
(1,4)-α-l-fuc, and (1,3)-β-d-gal |
317.5/8.5 |
1.1 |
18.6 |
At C2 and C4 (fuc and gal) |
[72] |
|
Spatoglossum schroederi |
Wild |
Gal, fuc, xyl, glcAc; Gal, fuc, xyl; |
(1,4)-β-d-gal, (1,4)-α-l-fuc, and (1,4)-β-d-xyl |
21.5; 21.5–24 |
0.5; 0.5 |
19; 2.1–2.9 * |
At C3 (gal) and C4 (fuc) |
|
|
Ectocarpales |
||||||||
|
Adenocystis utricularis |
Wild |
Gal, fuc, rha, man; Gal, fuc, rha; Gal, fuc, man |
(1,3)-α-l-fuc |
>100 |
5.53; 4.82; 5.53 |
23; 24; 23 |
At C4 (fuc and gal) |
|
|
Scytosiphon lomentaria |
Wild |
Gal, fuc, rha, xyl, man, uronic acid |
(1,3)-α-l-fuc, and (1,6)-β-d-gal |
8.5 |
7.33 |
29.5 |
At C3 and C4 (fuc), and C3 (gal) |
[78] |
|
Fucales |
||||||||
|
Cystoseira compressa |
Wild |
Gal, fuc |
(1,3)- and (1,4)-α-l-fuc |
100 |
2.32 |
14.7 |
At C2 and C4 (fuc) |
[64] |
|
Sargassum duplicatum |
Wild |
Gal, fuc |
(1,4)-α-l-fuc and β-d-gal (alternating) |
34–191 |
1 |
31.7 |
ND |
[14] |
|
S. feldmannii |
Wild |
Gal, fuc |
(1,3)-α-l-fuc |
183–184 |
2–2.6 |
25.3–32 |
At C2, C3 and C4 (fuc), and C2, C3, C4 and C6 (gal) |
|
|
S. fusiforme |
Wild |
Gal, fuc, xyl, Glc, glcAc, man, uronic acid; Gal, fuc, xyl, man, rha, glcAc, Glc |
(1,3)- and (1,4)-α-l-fuc |
90; 118.3/3.9 |
2; 3.7 |
17.5; 28.5 |
At C3 (fuc) |
|
|
S. hemiphyllum |
Wild |
Gal, fuc |
(1,6)-β-d-gal, (1,3)- and (1,4)-α-l-fuc, and (1,3)-β-d-gal |
148 |
4.5 |
32 |
At C2 and C4 (fuc) |
[82] |
|
S. mcclurei |
Wild |
Gal, fuc; Gal, fuc, man, xyl, glc |
(1,3)-α-l-fuc |
ND |
1.4; 2 |
35; 30.5 |
At C2 and C4 (fuc) |
|
|
S. patens |
Wild |
Gal, fuc, man, xyl, Glc, galactosamine |
ND |
424 |
1.9 |
14.4 |
ND |
|
|
S. polycystum |
Wild |
Gal, fuc, glc; Gal, fuc, man, xyl, glc |
(1,3)-α-l-fuc, and (1,6)-β-d-gal |
39.5; ND |
5.84; 1.48 |
33.6; 23.4 |
At C2 and C4 (fuc) |
|
|
S. siliquosum |
Wild |
Gal, fuc, glc, xyl, man, rha; Gal, fuc, Glc, xyl, man, rha, uronic acid |
(1,3)- and (1,4)-α-l-fuc |
107.3; ND |
1.9; 1.9 |
19.5; 20 |
At C4 and C6 (gal) |
|
|
S. thunbergii |
Wild |
Gal, fuc |
(1,3)-α-l-fuc |
7.2–333.5 |
5.26–5.88 |
27.2–30.1 |
At C2 and C4 (fuc), and C4 (gal) |
|
|
S. thunbergii |
Purchased from local store |
Gal, fuc |
(1,4)-α-d-gal, and (1,3)-β-l-fuc |
373 |
1.2 |
ND |
NA |
[90] |
|
S. wightii |
Wild |
Gal, fuc, Glc, man; Gal, fuc |
(1,3)-α-l-fuc |
>3.5; ND |
0.6; 3–3.5 |
379.1 †; 8.1–19.5 |
At C2 and/or C4 (fuc), or C2 and C3 (gal) |
|
|
Turbinaria ornata |
Wild |
Gal, fuc; Gal, fuc, man, xyl, glc |
(1,3)-α-l-fuc |
ND |
5; 1.2 |
32; 25.6 |
At C2 and/or C4 (fuc), and/or C2, C3, C4/C6 (gal) |
|
|
Laminariales |
||||||||
|
Alaria angusta |
Wild |
Gal, fuc |
(1,3)-α-l-fuc |
ND |
1.1 |
24 |
At C2 (fuc), and C2 and C4 (gal) |
[95] |
|
Costaria costata |
Wild |
Gal, fuc, man, rha, xyl |
ND |
ND |
1.2 |
18.9 |
ND |
[96] |
|
Ecklonia cava |
Wild |
Gal, fuc, man, rha; Gal, fuc, rha, glc |
ND |
ND |
4.8; 3.6 |
19.1; 22.2 |
At C2 (fuc) |
[96] |
|
Laminaria hyperborea |
ND |
Gal, fuc |
(1,3)-α-l-fuc |
469 |
44.5 |
53.8 |
At C2 and C4 (fuc) |
[12] |
|
Saccharina angustata |
Wild |
Gal, fuc, xyl, uronic acid |
(1,3)-, (1,4) and (1,2)-α-l-fuc |
56 |
9.1 |
4.2 |
At C4 (fuc and gal) |
[97] |
|
S. gurjanovae |
Wild |
Gal, fuc |
(1,3)-α-l-fuc |
123 |
3.2 |
25.1 |
At C2 and C4 (fuc), and C2 and/or C3 (gal) |
[98] |
|
S. japonica |
Wild |
Gal, fuc; Gal, fuc, man, xyl; Gal, fuc, man, rham, xyl; Gal, fuc, uronic acid, man, glcAc; Gal, fuc, Glc, man, rha, xyl; Gal, fuc, xyl, Glc, glcAc, rha, uronic acid |
(1,3)-α-l-fuc |
195/13.7; 1800; ND; 106.3; 23.5; 11 |
3.6; 1.1; 1.8; 9.1; 0.5; 10 |
21; 23.3; 23; 36.9; 18; 41.3 |
At C2 and C2/C4 (fuc) |
|
|
S. japonica |
Cultivated |
Gal, fuc; Gal, fuc, man, rham, xyl, Glc; Gal, fuc, man, Glc, rha, xyl, uronic acid |
(1,3)- and (1,4)-α-l-fuc |
261.7; 131.5; 8.1 |
3.8; 2.1; 5.8 |
11.4; 9.1; 41.8 |
At C4 (fuc) |
|
|
S. japonica |
Provided by Fujian Yida Food Co. |
Gal, fuc, man |
ND |
527.3 |
0.9 |
26.7 |
ND |
[105] |
|
S. japonica |
ND |
Gal, fuc |
(1,3)-α-l-fuc, and (1,6)-β-d-gal |
>10 |
3.5 |
48.3 |
At C4 and/or C2/C4 (fuc), and C4 and/or C3/C4 (gal) |
[106] |
|
S. latissima |
Wild |
Gal, fuc; Gal, fuc, xyl, man, Glc |
(1,3)-α-l-fuc |
416–449; 453 |
7.8; 4.1 |
0.8 ‡; 0.6 ‡ |
ND |
|
|
S. longicruris |
Wild |
Gal, fuc, xyl, man, Glc, glcAc; Gal, fuc, xyl, man, Glc, galAc, glcAc |
1529; 638 |
0.8; 0.4 |
17.6; 19.1 |
At C4 (fuc), and C3 (gal) |
[109] |
|
|
Undaria pinnatifida |
Wild |
Gal, fuc, man; Gal, fuc, rha; Gal, fuc, Glc, man, rha, xyl, ara |
(1,3)- or (1,4)-α-l-fuc |
ND; 290; ND |
1.1; 1.2; 1.3 |
29; 0.94 ‡; ND |
At C2, C3, C4 (fuc), or C2 and C4 (fuc and/or gal) |
|
|
U. pinnatifida (sporophylls) |
Wild |
Gal, fuc, xyl, man |
(1,3)-α-l-fuc |
>150 |
1.5 |
15 |
ND |
|
|
Cultivated |
Gal, fuc; Gal, fuc, man; Gal, fuc, xyl, man; Gal, fuc, man, xyl, uronic acid |
(1,3)-α-l-fuc, and (1,3)-, (1,4)-, (1,6)-β-d-gal |
ND; 1.4–3.7; 1246; 2100 |
1.4; 1.1; 1.1; 5 |
31; 8.4; 9.2; 7.4 |
At C2/C4 (fuc), and C3/C6 (gal) |
||
|
From mussel farms |
Gal, fuc, xyl, Glc, man; Gal, fuc, xyl, Glc, man, uronic acid |
171; >150 |
1.5; 1.5 |
15; 15 |
ND |
|||
|
U. pinnatifida |
From Marine Resources Pty Ltd. |
ND |
ND |
ND |
ND |
[118] |
||
|
From Marinova Pty Ltd. |
Gal, fuc, xyl, man |
(1,3)-α-l-fuc |
51.7 |
1.3 |
21.5 |
At C2 and C4 (fuc) |
[119] |
|
|
ND |
ND |
ND |
ND |
ND |
[120] |
|||
|
U. pinnatifida (sporophylls) |
ND |
Gal, fuc; Gal, fuc, uronic acid; Gal, fuc, xyl, man |
(1,3)-α-l-fuc, and (1,3)-, (1,4)-, (1,6)-β-d-gal |
9; 9; 104.4 |
0.9; 0.9; ND |
10.4; 10.4; 21 |
At C2 (fuc), and C3 and C6 (gal) |
|
|
Sphacelariales |
||||||||
|
Sphacelaria indica |
Wild |
Gal, fuc, xyl, man, Glc |
(1,3)-α-l-fuc |
26 |
3.3 |
4 |
At C4 (fuc) |
[124] |
ND, not detailed; NA, not applicable; * reported as molar ratio to fucose; † reported as mg/g fucoidan; ‡ reported as degree of sulfation.
3. Potential Pharmacological Activities
3.1. Anticancer/Antitumor Activity
Several studies have reported the anticancer/antitumor activities of galactofucans in different cancer cell lines, as well as antiproliferative, antimetastasis, and antiangiogenic effects (Table 2).
Table 2. G-fucoidans showing anticancer/antitumor activity with their respective sources and half-maximal inhibitory concentrations (IC50). Comparisons with standard or commercial compounds are also shown.
|
Source |
IC50 |
Compared with Standard/Commercial Compounds? |
References |
|---|---|---|---|
|
Saccharina latissima |
0.35 µg/mL (elastase inhibition) |
Yes. Superior to commercial heparins (UFH and tinzaparin) |
[107] |
|
Sargassum polycystum |
84.63 µg/mL (leukemia cells) and 93.62 µg/mL (breast cancer cells) |
No |
|
|
S. thunbergii |
29.7–93.5 μg/mL (inhibition of FGF1 binding) and 4.0–6.8 μg/mL (inhibition of FGF7 binding) |
No |
|
|
Undaria pinnatifida (sporophylls) |
0.10 mg/mL (breast adenocarcinoma) and 0.15 mg/mL (lung carcinoma) |
Yes. Superior to commercial fucoidan from Fucus for both cancer cell lines |
3.2. Antiviral Activity
|
Source |
EC50/IC50 |
Compared with Antiviral Drugs? |
References |
|---|---|---|---|
|
Adenocystis utricularis |
0.6–0.9 µg/mL (HIV-1) |
Yes. Superior to azidothymidine |
|
|
0.3 µg/mL (HSV-1) and 0.5 µg/mL (HSV-2) |
No |
[48] |
|
|
Dictyota dichotoma |
7.5 µg/mL (HSV-1), and 15.6 µg/mL (CVB3) |
Yes. Superior to ribavirin |
[68] |
|
Saccharina japonica |
0.001–0.005 µg/mL (HIV-1) |
No |
[100] |
|
0.2–25 µg/mL (HSV-1) |
Yes. Inferior to acyclovir and similar to heparin |
[97] |
|
|
Sargassum mcclurei |
0.96 µg/mL (HIV-1) |
Yes. Inferior to AMD3100 (plerixafor) |
[84] |
|
S. patens |
1.3 µg/mL (HSV-2), 5.5 µg/mL (HSV-1), and 4.1 µg/mL (HSV-1 acyclovir-resistant strain) |
No |
|
|
>50 µg/mL (virucidal activity against HSV-2), 1.3–1.65 µg/mL (plaque formation), 1.85–3.5 µg/mL (inhibition of virus adsorption) |
No |
||
|
1.5–5.5 mg/mL (HSV-1 replication) and 3–4 mg/mL (HSV-1 adsorption) |
Yes. Similar to acyclovir |
||
|
S. polycystum |
0.34 µg/mL (HIV-1) |
Yes. Inferior to AMD3100 (plerixafor) |
[84] |
|
Scytosiphon lomentaria |
0.76 µg/mL (HSV-1) and 1.34 µg/mL (HSV-2) |
No |
[78] |
|
Sphacelaria indica |
1.3 µg/mL (HSV-1) |
Yes. Superior to acyclovir when added to the overlay medium after penetration of the viruses into the host cell |
[124] |
|
Turbinaria ornata |
0.39 µg/mL (HIV-1) |
Yes. Inferior to AMD3100 (plerixafor) |
[84] |
|
Undaria pinnatifida |
0.77 µg/mL (HSV-1) |
Yes. Superior to acyclovir |
[111] |
|
32 µg/mL (HSV-1) and 0.5 µg/mL (HSV-2) |
Yes. Superior to acyclovir |
[120] |
|
|
U. pinnatifida (sporophylls) |
2.5 µg/mL (HSV-1), 2.6 µg/mL (HSV-2), and 1.5 µg/mL (HCMV) |
No |
|
|
U. pinnatifida |
1.1 µg/mL (HSV-1), 0.1 µg/mL (HSV-2), and 0.5 µg/mL (HCMV) |
No |
|
|
3.1 µg/mL (HSV-1) and 1.6 µg/mL (HSV-2) |
No |
[118] |
3.3. Anti-Inflammatory, Immunomodulatory, and Anticomplement Activities
3.4. Anticoagulant and Antithrombotic Activities
3.5. Antioxidant Activity
|
Source |
EC50/IC50 |
Compared with Standard/Commercial Compounds? |
References |
|---|---|---|---|
|
Cystoseira compressa |
0.43 mg/mL (DPPH) |
Yes. Inferior to ascorbic acid and butylated hydroxyanisole |
[64] |
|
Sargassum siliquosum |
2.58 mg/mL (DPPH) |
No |
[10] |
|
S. thunbergii |
0.22 mg/mL (superoxide radical), and 0.88 mg/mL (hydroxyl radical) |
Yes. Similar (hydroxy radical) or superior (superoxide radical) to vitamin C |
[90] |
3.6. Other Biological Activities
This entry is adapted from the peer-reviewed paper 10.3390/md20070412