Leucaena leucocephala (Lam.) de Wit is native to southern Mexico and Central America and is now naturalized in more than 130 countries. The spread of L. leucocephala is probably due to its multipurpose use such as fodder, timber, paper pulp, shade trees, and soil amendment. However, the species is listed in the world’s 100 worst invasive alien species, and an aggressive colonizer. It forms dense monospecific stands and threatens native plant communities, especially in oceanic islands. Phytotoxic chemical interactions such as allelopathy have been reported to play an important role in the invasion of several invasive plant species.
- invasive plant
- allelochemical
- mimosine
- phytotoxicity
- rhizosphere soil
1. Introduction

2. Allelopathy
2.1. Plant Extract
2.2. Leachate
2.3. Plant Litter and Residue
2.4. Rhizosphere Soil and Root Exudate
3. Allelochemical
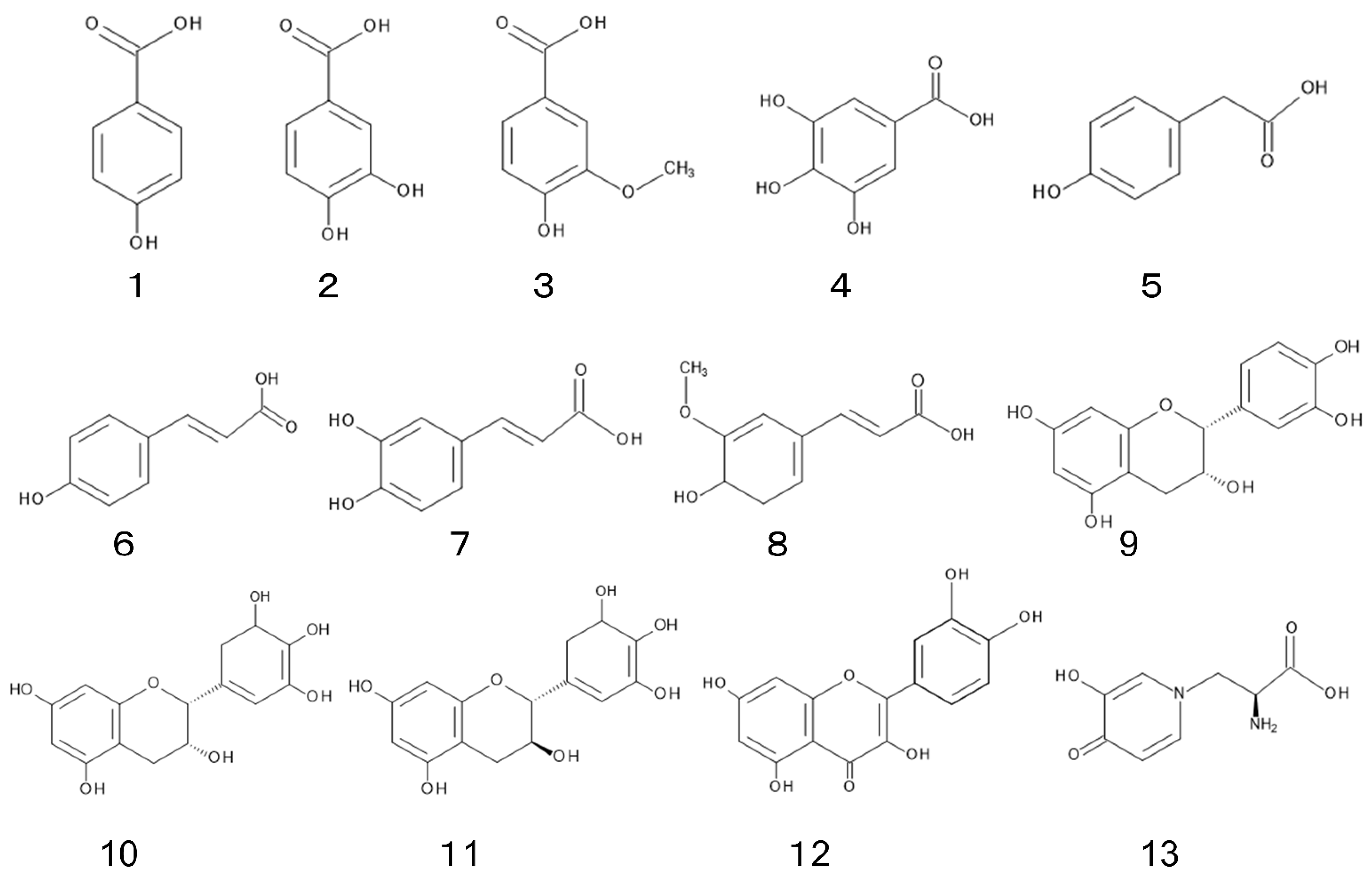
3.1. Phenolic Acid
Phenolic acids such as p-hydroxybenzoic acid (1), protocatechuic acid (2), vanillic acid (3), gallic acid (4), p-hydroxyphenylacetic acid (5), p-hydroxycinnamic acid (6), caffeic acid (7), and ferulic acid (8) were identified in the leaves of L. leucocephala. Total concentrations of those phenolic acids in young leaves were 2-fold greater than those in mature leaves [31]. The concentration of total phenolic compounds in L. leucocephala plants was estimated to be 1.3 to 2.8 mg g−1 of dry weight of the plants [43]. Phenolic acids have been found in a wide range of plants, plant residues, and soils, and their involvement has been often mentioned in the allelopathy of those plant species [44][45].
3.2. Flavonoid
3.3. Mimosine
4. Conclusions
This entry is adapted from the peer-reviewed paper 10.3390/plants11131672
References
- Invasive Species Compendium. Leucaena leucocephala (Leucaena). Available online: https://www.cabi.org/ISC/abstract/20127801953 (accessed on 8 April 2022).
- Global Invasive Species Database. Species Profile: Leucaena leucocephala. Available online: http://www.iucngisd.org/gisd/speciesname/Leucaena+leucocephala (accessed on 8 April 2022).
- National Research Council. Leucaena: Promising Forage and Tree Crop for the Tropics; The National Academies Press: Washington, DC, USA, 1984; pp. 1–129.
- Van den Beldt, R.J.; Brewbaker, J.L. Leucaena Wood Production and Use; Nitrogen Fixing Tree Association: Waimanalo, HI, USA, 1985; pp. 1–50.
- Verma, S. A review study on Leucaena leucocephala: A multipurpose tree. Int. J. Sci. Res. Sci. Eng. Technol. 2016, 2, 103–105.
- Abair, A.; Hughes, C.E.; Bailey, C.D. The evolutionary history of Leucaena: Recent research, new genomic resources and future directions. Trop. Grassl. Forrajes Trop. 2019, 7, 65–73.
- Sohtome, Y.; Tokunaga, T.; Ueda, K.; Yamamura, S.; Ueda, M. Leaf-closing substance in Leucaena leucocephala. Biosci. Biotechnol. Biochem. 2002, 66, 51–56.
- Walton, C. Leucaena (Leucaena leucocephala) in Queensland; Department of Natural Resources and Mines: Brisbane, QLD, Australia, 2003; pp. 1–51.
- Olckers, T. Biological control of Leucaena leucocephala (Lam.) de Wit (Fabaceae) in South Africa: A tale of opportunism, seed feeders and unanswered questions. Afr. Entomol. 2011, 19, 356–365.
- Brewbaker, J.L.; Hegde, N.; Hutton, E.M.; Jones, R.J.; Lowry, J.B.; Moog, F.; van den Beldt, R. Leucaena—Forage Production and Use; Nitrogen Fixing Tree Association: Waimanalo, HI, USA, 1985; pp. 1–39.
- Campbell, S.; Vogler, W.; Brazier, D.; Vitelli, J.; Brooks, S. Weed Leucaena and its significance, implications and control. Trop. Grassl. Forrajes Trop. 2019, 7, 280–289.
- Garcia, G.W.; Ferguson, T.U.; Neckels, F.A.; Archibald, K.A.E. The nutritive value and forage productivity of Leucaena leucocephala. Anim. Feed Sci. Technol. 1996, 60, 29–41.
- Wanapat, M.; Kang, S.; Polyorach, S. Development of feeding systems and strategies of supplementation to enhance rumen fermentation and ruminant production in the tropics. J. Anim. Sci. Biotechnol. 2013, 4, 32.
- De Angelis, A.; Gasco, L.; Parisi, G.; Danieli, P.P. A multipurpose Leguminous plant for the mediterranean countries: Leucaena leucocephala as an alternative protein source: A review. Animals 2021, 11, 2230.
- Durango, S.G.; Barahona, R.; Bolívar, D.; Chirinda, N.; Arango, J. Feeding strategies to increase nitrogen retention and improve rumen fermentation and rumen imcrobial population in beef steers fed with tropical forages. Sustainability 2021, 13, 10312.
- Anupam, K.; Swaroop, V.; Deepika, L.P.S.; Bist, V. Turning Leucaena leucocephala bark to biochar for soil application via statistical modelling and optimization technique. Ecol. Eng. 2015, 82, 26–39.
- Jha, P.; Neenu, S.; Rashmi, I.; Meena, B.P.; Jatav, R.C.; Lakaria, B.L.; Biswas, A.L.; Singh, M.; Patra, A.K. Ameliorating effects of Leucaena biochar on soil acidity and exchangeable Ions. Commun. Soil Sci. Plant Anal. 2016, 47, 1252–1262.
- Dalzell, S.A. Leucaena cultivars—Current releases and future opportunities. Trop. Grass. 2019, 7, 56–64.
- Khanna, N.K.; Shukla, O.P.; Gogate, M.G.; Narkhede, S.L. Leucaena for paper industry in Gujarat, India: Case study. Trop. Grassl. Forrajes Trop. 2019, 7, 200–209.
- Mac Dicken, K.G. Nitrogen Fixing Trees for Wastelands; FAO Regional Office for Asia and the Pacific—Food and Agriculture Organization of the United Nations: Bangkok, Thailand, 1988; pp. 1–104.
- Hansted, A.L.S.; Nakashima, G.T.; Martins, M.P.; Yamamoto, H.; Yamaji, F.M. Comparative analyses of fast growing species in different moisture content for high quality solid fuel production. Fuel 2016, 184, 180–184.
- Nguyen, M.P.; Vaast, P.; Pagella, T.; Sinclair, F. Local knowledge about ecosystem services provided by trees in coffee agroforestry practices in northwest Vietnam. Land 2020, 9, 486.
- Yu, G.; Huang, H.Q.; Wang, Z.; Brierley, G.; Zhang, K. Rehabilitation of a debris-flow prone mountain stream in southwestern China—Strategies, effects and implications. J. Hydrol. 2012, 414–415, 231–243.
- Adhikary, P.P.; Hombegowda, H.C.; Barman, D.; Jakhar, P.; Madhu, M. Soil erosion control and carbon sequestration in shifting cultivated degraded highlands of eastern India: Performance of two contour hedgerow systems. Agrof. Syst. 2017, 91, 757–771.
- Ishihara, K.L.; Honda, M.D.H.; Bageel, A.; Borthakur, D. Leucaena leucocephala: A leguminous tree suitable for eroded habitats of Hawaiian Islands. In Ravine Lands: Greening for Livelihood and Environmental Security; Dagar, J., Singh, A., Eds.; Springer: Singapore, 2018; pp. 413–431.
- Standley, P.C. Trees and Shrubs of Mexico, Contributions from the United State National Herbarium; Government Printing Office: Washington, DC, USA, 1922; pp. 1–1721.
- Rice, E.L. Allelopathy, 2nd ed.; Academic Press: Orlando, FL, USA, 1984; pp. 1–422.
- Bais, H.P.; Weir, T.L.; Perry, L.G.; Gilroy, S.; Vivanco, J.M. The role of root exudates in rhizosphere interactions with plants and other organisms. Annu. Rev. Plant Biol. 2006, 57, 233–266.
- Bonanomi, G.; Sicurezza, M.G.; Caporaso, S.; Esposito, A.; Mazzoleni, S. Phytotoxicity dynamics of decaying plant materials. New Phytol. 2006, 169, 571–578.
- Belz, R.G. Allelopathy in crop/weed interactions—An update. Pest. Manag. Sci. 2007, 63, 308–326.
- Chou, C.H.; Kuo, Y.L. Allelopathic research of subtropical vegetation in Taiwan. J. Chem. Ecol. 1986, 12, 1431–1448.
- Boaprem, P. Allelopathic effects of leucaena leaves extract (Leucaena leucocephala (Lam.) de Wit) on the growth of rice (Oryza sativa L.), wrinkle duck-beak (Ischaemum rugosum Salisb), and mung bean (Vigna radiata (L.) R. Wilczek). Songklanakarin. J. Sci. Technol. 2019, 41, 619–623.
- Ishak, M.S.; Sahid, I. Allelopathic effects of the aqueous extract of the leaf and seed of Leucaena leucocephala on three selected weed species. AIP Conf. Proc. 2016, 1744, 020029.
- Sahoo, U.K.; Upadhyaya, K.; Meitei, C.B. Allelopathic effects of Leucaena leucocephala and Tectona grandis on germination and growth of maize. Allelopath. J. 2007, 20, 135–144.
- John, J.; Sreekumar, K.M.; Rekha, P. Allelopathic effects of leaf leachates of multipurpose trees on vegetables. Allelopath. J. 2007, 19, 507–516.
- Maiti, P.P.; Bhakat, R.K.; Bhattacharjee, A. Allelopathic effects of Lantana camara on physio-biochemical parameters of Mimosa pudica seeds. Allelopath. J. 2008, 22, 59–67.
- Kalpana, P.; Navin, M.K. Assessment of allelopathic potential of Leucaena leucocephala (Lam) De Vit on Raphanus sativus L. Int. J. Sci. Res. Publ. 2015, 5, 1–3.
- Chai, T.T.; Ooh, K.F.; Ooi, P.W.; Chue, P.S.; Wong, F.C. Leucaena leucocephala leachate compromised membrane integrity, respiration and antioxidative defense of water hyacinth leaf tissues. Bot. Stud. 2013, 54, 8.
- Ahmed, T.; Hoque, A.T.M.R.; Hossain, M.K. Allelopathic effects of Leucaena leucocephala leaf litter on some forest and agricultural crops grown in nursery. J. For. Res. 2008, 19, 298–302.
- Ishak, M.S.; Ismail, B.S.; Yusoff, N. Allelopathic potential of Leucaena leucocephala (Lam.) de Wit on the germination and seedling growth of Ageratum conyzoides L., Tridax procumbens L. and Emilia sonchifolia (L.) DC. Allelopath. J. 2016, 37, 109–122.
- Kamara, A.Y.; Akobundu, I.O.; Sanginga, N.; Jutzi, S.C. Effects of mulch from 14 multipurpose tree species (MPTs) on early growth and nodulation of cowpea (Vigna unguiculata L.). J. Agron. Crop Sci. 1999, 182, 127–133.
- Parvin, R.; Shapla, T.L.; Amin, M.H.A. Effect of leafs and Leucaena leucocephala different three depth soil on the allelopathy of agricultural crops. J. Innov. Dev. Strategy 2011, 5, 61–69.
- Sharma, P.; Chaurasia, S. Evaluation of total phenolic, flavonoid contents and antioxidant activity of Acokanthera oppositifolia and Leucaena leucocephala. Int. J. Pharmacogn. Phytochem. Res. 2014, 15, 175–180.
- Inderjit. Plant phenolics in allelopathy. Bot. Rev. 1996, 62, 186–202.
- Dalton, B.R. The occurrence and behavior of plant phenolic acids in soil environments and their potential involvement in allelochemical interference interactions: Methodological limitations in establishing conclusive proof of allelopathy. In Principals and Practices in Plant Ecology: Allelochemical Interactions; Inderjit, Dakshini, K.M.M., Foy, C.L., Eds.; CRC Press: Boca Raton, FL, USA, 1999; pp. 57–74.
- Erickson, A.J.; Ramsewak, R.S.; Smucker, A.J.; Nair, M.G. Nitrification Inhibitors from the Roots of Leucaena leucocephala. J. Agric. Food Chem. 2000, 48, 6174–6177.
- Xu, Y.; Tao, Z.; Jin, Y.; Yuan, Y.; Dong, T.T.X.; Tsim, K.W.K.; Zhou, Z. Flavonoids, a potential new insight of Leucaena leucocephala foliage in ruminant health. J. Agric. Food Chem. 2018, 66, 7616–7626.
- Barbehenn, R.V.; Constabel, C.P. Tannins in plant–herbivore interactions. Phytochemistry 2011, 72, 1551–1565.
- Zhou, M.; Wei, L.; Sun, Z.; Gao, L.; Meng, Y.; Tang, Y.; Wu, Y. Production and transcriptional regulation of proanthocyanidin biosynthesis in forage legumes. Appl. Microbiol Biotechnol. 2015, 99, 3797–3806.
- Paengkoum, S.; Petlum, A.; Purba, R.A.P.; Paengkoum, P. Protein binding affinity of various condensed tannin molecular weights from tropical leaf peel. J. Appl. Pharm. Sci. 2021, 11, 114–120.
- Renz, J. Mimosine. Physiol. Chem. 1936, 244, 153–158.
- Brewbaker, J.L.; Hylin, J.W. Variations in mimosine content among Leucaena species and related mimosaceae. Crop Sci. 1965, 5, 348–349.
- Smith, I.K.; Fowden, L.A. Study of mimosine toxicity in plants. J. Exp. Bot. 1996, 17, 750–761.
- Soedarjo, M.; Borthakur, D. Mimosine produced by the tree-legume Leucaena provides growth advantages to some Rhizobium strains that utilize it as a source of carbon and nitrogen. Plant Soil 1996, 186, 87–92.
- Nguyen, B.C.; Tawata, S. The chemistry and biological activities of mimosine: A review. Phytother. Res. 2016, 30, 1230–1242.
- John, J.; Narwal, S.S. Allelopathic plants. 9. Leucaena Leucocephala (Lam.) De Wit. Allelopath. J. 2003, 12, 13–36.
- Hussain, M.I.; Gonzalez-Rodriguez, L.; Roger, M.R. Germination and growth response of four plant species to different allelochemicals and herbicides. Allelopath. J. 2008, 22, 101–110.