Multiple sclerosis (MS) is a chronic inflammatory disease of the central nervous system and its pathophysiology is characterized by a progressive blood-brain barrier dysfunction accompanied by infiltration in the central nervous system of peripheral pathogenic immune cells and inflammatory mediators leading to demyelination, axonal damage, and dysfunction and/or loss of synapses. Accumulating evidence highlights blood and cerebrospinal fluid (CSF) derived extracellular vesicles (EVs) as potential biomarkers of MS disease stages and of response to treatment. In particular, EVs released from blood–brain barrier (BBB) endothelial cells, platelets, leukocytes, myeloid cells, astrocytes, and oligodendrocytes seem to be involved in the pathogenesis of MS and of its rodent model experimental autoimmune encephalomyelitis. Further research is necessary to validate these observations and the screening of specific EVs subsets based on their cargo and membrane compositions associated to specific MS pathophysiological mechanisms might help guiding MS diagnosis, prognosis, and response to therapy.
- Multiple Sclerosis
- Extracellular Vesicles
- Biomarkers
- Neuroinflammation
- cerebrospinal fluid
- MS therapy
- experimental autoimmune encephalomyelitis
Dear author, the following contents are excerpts from your papers. They are editable.
1. Multiple Sclerosis: Clinical Features and Pathophysiology
2. Extracellular Vesicles as Potential Biomarkers in MS
| EVs Cellular Origin | Surface Marker | Functional Implication | Detection Levels | References | Study Size |
|---|---|---|---|---|---|
| Serum/Plasma | |||||
| Endothelial cells | CD31+ | Acute BBB disruption and contribution in Gad+ MRI active lesions |
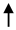 E-MS RRMS E-MS RRMS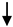 R-MS; SPMS; HC R-MS; SPMS; HC |
Minagar et al., 2001 [40] | 48 HC; 30 E-RRMS; 20 R-RRMS |
| Jy et al., 2004 [41] | 35 HC; 30 E-RRMS; 20 R-RRMS | ||||
| Alexander et al., 2015 [42] | 36 HC; 44 RRMS; 16 SPMS |
||||
| CD4+ and CD8+ T-lymphocytes activation | Wheway et al., 2014 [43] | ||||
| CD51+ | Chronic endothelial injury | E-MS; R-MS HC |
Minagar et al., 2001 [40] | 48 HC; 30 E-RRMS; 20 R-RRMS | |
| CD54+ CD62E CD106+ |
Monocytes conjugates for endothelial adhesion | E-MS R-MS; HC |
Jy et al., 2004 [41] | 35 HC; 30 E-RRMS; 20 R-RRMS | |
| Jimenez et al., 2005 [44] | 10 HC; 11 E-RRMS; 9 R-RRMS |
||||
| Monocytes | CD14+ | Acute endothelial injury | R-RRMS HC and SPMS |
Saenz-Cuesta et al., 2014 [45] | 20 HC; 13 SPMS 64 R-RRMS |
| Leukocytes | CD45+ | Acute endothelial injury | R-RRMS HC and SPMS |
Saenz-Cuesta et al., 2014 [45] | 20 HC; 13 SPMS 64 R-RRMS |
| Platelets | CD62p CD41+/ CD61+ |
Platelets activation and leukocytes interaction with damaged endothelium | R-MS (RRMS) HC and SPMS |
Saenz-Cuesta et al., 2014 [45] | 20 HC; 13 SPMS 64 R-RRMS |
| Sheremata et al., 2008 [46] | 92 HC; 33 R-RRMS | ||||
| CD42b+ | Incremented experimental BBB permeability (except for CIS) |
MS (PMS; RMS; CIS) HC |
Marcos-Ramiro et al., 2014 [47] | 49 HC; 23 SPMS; 51 RRMS; 12 CIS; 9 PPMS |
|
| Cerebrospinal Fluid | |||||
| Microglia/Macrophage | IB-4 | Acute BBB disruption and contribution in Gad+ MRI active lesions |
E-MS (CIS, RRMS) R-MS (RRMS); HC |
Verderio et al., 2012 [16] | 13 HC; 39 R- RRMS 28 E-RRMS; 28 CIS |
| T-cells | CCR3/ CCR5 CD4/ CCR3 CD4/ CCR5 |
Acute BBB disruption and contribution in Gad+ MRI active lesions |
E-MS (RRMS) R-MS (RRMS) |
Geraci et al., 2018 [48] | 10 R-RRMS; 13 E-RRMS |
2.1. Serum and Plasma
| miRNA | Detection Level | Study Size | Functional Implication | Reference |
|---|---|---|---|---|
| miR-15b-5p | In RRMS vs. HC |
14 RRMS 11 S/PPMS 11 HC |
Targets FGF-2 implicated in demyelination and remyelination | Ebrahimkhani et al., 2017 [51] |
| miR-451a | In RRMS vs. HC |
14 RRMS 11 S/PPMS 11 HC |
Regulator of oxidative stress | Ebrahimkhani et al., 2017 [51] |
| miR-30b-5p | In RRMS vs. HC |
14 RRMS 11 S/PPMS 11 HC |
Neuro-axonal injury | Ebrahimkhani et al., 2017 [51] |
| miR-342-3p | In RRMS vs. HC |
14 RRMS 11 S/PPMS 11 HC |
Neuro-axonal injury | Ebrahimkhani et al., 2017 [51] |
| miR-127-3p | In S/PPMS vs. HC |
14 RRMS 11 S/PPMS 11 HC |
Ebrahimkhani et al., 2017 [51] | |
| miR-370-3p | In S/PPMS vs. HC |
14 RRMS 11 S/PPMS 11 HC |
Ebrahimkhani et al., 2017 [51] | |
| miR-409-3p | In S/PPMS vs. HC |
14 RRMS 11 S/PPMS 11 HC |
Ebrahimkhani et al., 2017 [51] | |
| miR-432-5p | In S/PPMS vs. HC |
14 RRMS 11 S/PPMS 11 HC |
Ebrahimkhani et al., 2017 [51] | |
| miR-122-5p | In remission RRMS vs. HC; in relapse RRMS vs. remission RRMS |
30 Remission- RRMS 33 Relapse- RRMS 32 HC |
Targets STAT3 and AHR (not validated), regulators of differentiation of Th17 and immunosuppressive T cells | Selmaj et al., 2017 [53] |
| miR-196b-5p | In relapse RRMS vs. HC; in relapse RRMS vs. remission RRMS |
30 Remission- RRMS 33 Relapse- RRMS 32 HC |
Targets STAT3 and AHR (not validated), regulators of differentiation of Th17 and immunosuppressive T cells | Selmaj et al., 2017 [53] |
| miR-301a-3p | In relapse RRMS vs. HC |
30 Remission- RRMS 33 Relapse- RRMS 32 HC |
Targets STAT3 and AHR (not validated), regulators of differentiation of Th17 and immunosuppressive T cells | Selmaj et al., 2017 [53] |
| miR-532-5p | In relapse RRMS vs. HC; in relapse RRMS vs. remission RRMS |
30 Remission- RRMS 33 Relapse- RRMS 32 HC |
Targets STAT3 and AHR (not validated), regulators of differentiation of Th17 and immunosuppressive T cells | Selmaj et al., 2017 [53] |
| Let-7i | In MS vs. HC |
4 MS 4 HC |
Inhibition of Treg cells differentiation from naive CD4+ T cells | Kimura et al., 2018 [54] |
| miR-146a-5p | Detected in CSF exosomes | MS = 10 | Synaptic alterations in in vitro experiments | Prada et al., 2018 [57] |
| miR-219a-5p | Artificially enriched exosomes | EAE mice | Maturation of OPCs; clinical score improvement | Osorio-Querejeta et al., 2020 [58] |
2.2. Cerebrospinal Fluid
2.3. EVs as a Biomarker of Pharmacological Response
This entry is adapted from the peer-reviewed paper 10.3390/ijms21197336