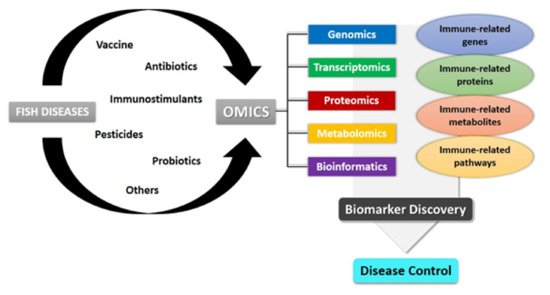
4. Multi-Omics Perspective on Infectious Fish Diseases Studies
The use of high-throughput omics approaches is becoming a powerful multidisciplinary tool for life science research, including fish disease studies. Omics approaches to study DNA variations (genomics), gene expressions at mRNA level (transcriptomics), protein expression (proteomics), and metabolite concentration (metabolomics) are essential for investigating the interplay between fish immune systems and pathogen infections [
92]. The basic aims of these omics are to provide a particular conception and understanding of complex biological systems as a whole [
93,
94], especially on the infected fish. Furthermore, these approaches allow identifying gene networks and molecular pathways associated with the fish immune response towards pathogens [
8]. Extensive applications of the multi-omics approach in fish disease studies could suggest multiple solutions to combat the increasing mortality of reared fishes on a cellular level as the current methods used, such as antibiotics and vaccinations are not sustainable in the long term.
Multi-omics or integration of omics data analysis provides a better understanding of molecular mechanisms of the fish immune system, regarding gene expression/regulation, protein expression, and metabolite production. The high-throughput data sets from different levels of omics studies can be used as a powerful method for the development of vaccines and immunostimulants that can be used in preventing and controlling disease infection in fish. The use of advanced technology such as next-generation sequencing (NGS), can directly be identified and specifically detect the presence of infection before any clinical signs appear in the fish [
95]. In proteomics study, a high-throughput technology has been often used to identify novel antigens for new vaccine development. A study by Pang et al. [
96], had identified outer membrane protein expression known as dihydrolipoamide dehydrogenase (DLD), to act as a vaccine candidate for
Epinephelus coioides against three
Vibrio species (
V. harveyi, V. alginolyticus, and V. parahaemolyticus). In another study, tilapias induced with exogenous l-leucine elevated the serine and proline to eliminate
Streptococcus iniae infection suggesting that the two metabolites play a crucial role in pathogen elimination by the host [
97]. Elucidation of the mechanisms by either genomic, proteomic, and metabolomics would benefit the idea of the development of a novel vaccine or immunostimulant method to boost fish immune response from bacterial invasions, which simultaneously reduce or solve the fish disease problems.
As this review aims to compile the vast literature on the multi-omics approaches including transcriptomics, proteomics, and metabolomics for advancements management in fish disease studies, we will first describe each of the omics applications individually to assess their relevance used in fish’s immune response studies and then walk the readers through the biomarkers found from these studies. Table 2 compiles the studies on fish disease using various omics approaches. As each of the omics approaches uses various methods to analyze the available data, the table also gathers the methods used in each of the studies.
4.1. Transcriptomics
The transcriptomic approach has been extensively applied in many fish disease studies such as in grass carp (
Ctenopharyngodon idellus) [
98], rainbow trout (
O. mykiss) [
99], tilapia (
O. niloticus) [
100], pacific cod (
Gadus microcephalus) [
101], orange-spotted grouper (
E. coioides) [
102], brown-marble grouper (
E. fuscoguttatus) [
103] and others (
Table 2). These studies have enabled the discovery of many immune-related genes and their involvement in immune-specific pathways that can eventually guide disease diagnosis and prevention. Previous studies conducted by [
102,
104] have utilized the RNA-seq technology to determine the transcriptional responses of orange-spotted groupers,
E. coioides to Singapore grouper iridovirus (SGIV),
V. alginolyticus, and
V. harveyi infection, respectively. These studies highlighted the discovery of genes with altered expression in response to the infection, such as complement component-related genes, hepcidin-like antimicrobial peptide precursor, lectin, interleukins, and interferon-related genes. These genes were previously reported to have a significant role in the fish immune system [
102,
104]. Hepcidin is well-known as one of the important antimicrobial peptides (AMPs) in fish. It is an essential mediator for the innate immune response as it is usually present at the fish mucosal barrier and plays a key role in the fish’s first line of defense [
102]. Hepcidin also catalyzes direct broad-spectrum antimicrobial activities, thus functioning in antimicrobial immune responses and iron homeostasis [
105]. Besides hepcidin, previous studies have shown the importance of lectins as the recognition molecules that facilitate innate and adaptive immune responses. Lectins are known for their ability to bind carbohydrates due to the presence of their carbohydrate-recognition domain (CRD), which enables them to function as pathogen-recognition receptors (PRRs) and recognize the carbohydrates of bacteria [
106,
107]. This eventually leads to the other immunological functions of lectins such as cell agglutination, antiviral activities, phagocytosis, encapsulation, and production of antimicrobial peptides [
106,
107,
108]. Interleukins and interferons are also important to the fish immune response and these groups of cytokines are mainly involved in inflammatory responses and antiviral defense, respectively [
109,
110,
111,
112].
Transcriptomic analysis of Asian seabass,
Lates calcarifer on post-infection by nervous necrosis virus (NNV) [
113] revealed an increase in the expression of genes involved in chemokine signaling pathways such as CC chemokines and CXC chemokines that are similar to a study in the liver of large yellow croaker,
Larimichthys corcea infected against
C. irritans [
114]. The chemokines family has been recognized for its importance in many biological processes including innate and adaptive responses. Functionally, chemokines are divided into inflammatory chemokines and homeostatic chemokines [
115]. The inflammatory chemokines such as CCL1-5 and CXXL1-11 generally regulate neutrophil and activated T-cell migration. Most CC chemokines are crucial for inducing chemotaxis monocytes, T-cells, and macrophages, leading to the innate immune response [
116]. Meanwhile, homeostatic chemokines such as CXCL12 are essential for the migration of antigen-presenting cells (APC) and lymphocytes to the lymph node where immune surveillance occurs. These chemokines are also important for the migration of T cells to the tissues with APC. These actions are important for effective adaptive immune responses [
116,
117].
Transcriptomics approach not only provides an insight into the response of the fish towards different types of infection at the transcriptome level but also helps in revealing the complexity of the host’s defense mechanism against infection [
103] by the identification of potential key resistance genes that is important to improve the immunity of fish for disease resistance. However, while transcriptome profiling can be comprehensive, it may be subjected to post-transcriptional and post-translational modifications. As such, proteomics is needed to verify and validate the protein levels.
4.2. Proteomics
Several reports have described the applications of mass spectrometry-based (MS-based) proteomics in studying the fish proteome response such as in Japanese flounder (
Paralichthys olivaceus) [
118], common carp (
Cyprinus carpio) [
119], Atlantic salmon [
120], and many more (
Table 2). A comparative analysis of protein expression in the spleen of healthy and infectious spleen and kidney necrosis virus (ISKNV) infected mandarin fish (
Siniperca chuatsi) revealed the up-regulated autophagy-related proteins including LC3 and P13Ks indicated the induction of CPB cells (Chinese perch brain cells) autophagy in the early-stage infection [
121]. Meanwhile, the heat-shock protein 27 (HSP27) family was found to be up-regulated in response to grouper nervous necrosis virus (RGNNV) infection in the sea perch [
122]. The up-regulation was similar to other fish-virus and bacterial studies [
123,
124], highlighting it as a therapeutic target for virus or bacteria-associated diseases. Furthermore, the proteomics approach of zebrafish skin infected with
Aeromonas hydrophila revealed the increased expression of actin, myosin heavy chain, and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) [
125]. The expression of both actin and myosin in the skin indicated that these proteins are important for the skin response of zebrafish to
A. hydrophila infection. A few studies have shown that actins play significant roles in resistance to bacterial or viral infections [
126]. The presence of actin such as F-actin and other small GTPases Rho, Rac, and cell division cycle 42 were reported to be important in cell motility of fish as it aids in initiating the phagocytic processes for invasion of pathogens via rearrangement of the actin cytoskeleton [
127,
128]. Myosin protein is composed of two heavy chains and two light chains with ATPase activity and actin-binding sites. It provides contractile force through hydrolysis of ATPs and interaction with actin [
129,
130]. Myosin was suggested in a previous study to be activated during an immune response. It was suggested that the increased disease resistance in fish might be associated with the low expression of myosin. A low expression of myosin was proposed to inhibit muscle growth, which inhibits further pathogen infection [
125]. Meanwhile, GAPDH was reported to have multiple roles in immune function, defense responses, antioxidant, and energy metabolism in fish [
131].
Other immune-related proteins expressed due to
V. anguillarum infection are mannan-binding lectin (MBL) and cyclophilins in the skin mucus of Atlantic cod,
Gadus morhua [
132]. Thus, these proteomics studies have shown the importance of an advanced high-throughput proteomic approach in disclosing the complication raised regarding fish response towards disease infection at the protein level. However, this alone will not provide sufficient information to wholly understand all the underlying mechanisms that play a role in responding to a pathogenic threat. Metabolomics study complements the two omics as mentioned earlier in this way as the slightest change in the environment will affect the metabolites produced by an organism. This particular advantage is utilized in the many metabolomics studies that are discussed below.
4.3. Metabolomics
Similar to the other omics, metabolomics has continued to grow rapidly and is considered a prominent tool that predicts and explains the complex phenotypes in a diverse biological system. It acts as a complementary platform and is often used in accordance with transcriptomics and proteomics since metabolomics is a downstream result of gene and protein expression. Metabolomics utilizes the high-throughput screening of various samples using instruments to assess any chemical risk. A study by Du et al. [
97] reported on the effects of l-leucine-induced metabolome to eliminate
Streptococcus iniae in tilapias, exploring the metabolomics approach’s potential in infectious diseases. Other studies on the metabolome changes during bacterial infection also have been conducted on several fish species such as tilapia [
133], Atlantic salmon [
40,
134], crucian carp [
135], and grouper [
136]. A recent study by Low et al. [
49], discussed the limitations and the challenges of metabolomics applications in fish disease studies while emphasizing the impacts of infectious fish diseases and the possibility of enhancing disease resistance in fish using metabolomics.
Activities of the cells are reflected by the metabolome changes that can identify complex biologically essential changes [
135]. In exploring targeted compounds and pathways necessary for crucian carps defense against infection caused by
Edwardsiella tarda, a gas chromatography–mass spectrometry-based (GC-MS-based) metabolomics approach was used in this research. The research has highlighted elevation in unsaturated fatty acid biosynthesis and decreased fructose and mannose metabolism. In contrast, the increase in palmitic acid and decrease in D-mannose were highlighted as the most crucial metabolic difference. These pathways and metabolites were suggested to be the central biomarkers in differentiating survivals from death in crucian carps infected by
E. tarda. Their findings stressed the importance of metabolic strategy in studying and understanding fish responses to bacterial infections. Thus, targeted compounds and pathways identified can be a good reference for further analysis on host-pathogen interaction.
In another study [
136], the high concentration of amino acid such as valine and leucine detected in the susceptible group of groupers infected with vibriosis is related to the activation of anti-bacterial infection pathways that involves valine, leucine, and isoleucine metabolism. Identification of these metabolites (valine and leucine) and their specific functions in inhibiting or escalating the immune response has enabled multiple other research to be conducted in order to improve the current problems in fish disease. The regulatory role of amino acids such as leucine, isoleucine, and valine are reported in various fish studies centered around identifying the dietary and regulatory effects of amino acids in fish and their impact on the immune system [
40]. In a study conducted by Castro et al. [
6] that aimed to observe the immune responses at an early stage in rainbow trout (
O. mykiss) infected with viral hemorrhagic septicemia virus, it was found that the liver is an immunocompetent organ that has a vital role in the immune response. Furthermore, a study conducted by Lardon et al. [
137] on common carps (
C. carpio) found that the liver contained metabolites associated with the general energy pathway such as ATP/ADP, phosphocreatine, lactate, and amino acids metabolites such as valine, (iso)leucine, alanine, glutamate, glutamine, glycine, and aspartate were significantly decreased in fish that were induced with hypoxia compared to control fish. The decreased of these metabolites was due to oxygen limitation availability, which minimize the consumption of ATP/ADP and simultaneously decreased the accumulation of lactate to conserve their energy consumption [
138]. As for amino acids, the decrease of the metabolites was due to the use of amino acids in regulating the growth and metabolism of the fish [
139]. While hypoxia is not an infectious disease, the lack of oxygen in the fish system is seen to achieve a similar stress response in the fish when infected with pathogens, according to the results of a study conducted by Lardon et al. [
140]. Since amino acids are vital to regulating key metabolic pathways related to maintenance and immune responses, identifying specific amino acids unique to certain biological pathways is essential to discover biologically active metabolites [
136,
137]. In another study, the effects of waterborne chlorpyrifos in freshwater carp,
Cyprinus carpio had caused the increased biosynthesis of valine, leucine, and isoleucine for oxidative/ stress metabolism, while the increased of, alanine and lactate pattern suggests that the subsequent metabolic pathway was inhibited. An increase of pyruvate is found to inhibit the metabolic pathway leading to the acetyl-CoA formation that will impact the tricarboxylic acid cycle (TCA cycle) and in turn the energy metabolism of the fish. As a large part of the energy is allocated to maintain and activate the immune system, inhibition of the TCA cycle will undoubtedly cause a delay in the immune response of the infected fish [
141].
The roles played by metabolites are again emphasized in a metabolomics analysis conducted by Liu et al. [
40] to profile the metabolites in the kidneys of Atlantic salmon infected with
A. salmonicida. It was found that the metabolites extracted from the infected samples have an altered profile compared to the control ones. Four pathways were reconstructed from the significantly altered metabolites (fumarate, alanine, valine, glycine, choline, glycerophosphorylcholine (GPC), and aspartate) that were obtained from the proton nuclear magnetic resonance (H-NMR) analysis. These metabolic pathways are the citrate cycle, glycolysis/gluconeogenesis, tryptophan metabolism, and the urea cycle. While alanine and glycine are found to be upregulated mid-experiment, valine shows the opposing result. While not directly affecting the innate immune system, these cycles play a role in producing the intermediates. This study by Liu et al. [
40] suggests that alanine and glycine might play a role in protein synthesis, benefiting the viral genome synthesis.
A study by Nurdalila et al. [
142] that involved metabolite fingerprinting of
E. fuscoguttatus infected with
V. vulnificus had fascinating findings where omega 9 (ω-9) was found to be a potential biomarker for vibriosis in brown marbled groupers. Regulation of immune cell function by short-chain fatty acids with the host immune response and pathogen resistance may be influenced by fatty acids that play a significant role as demonstrated by human and animal studies, during ex vivo and in vitro experiments [
143]. This study was essentially based on the speculation that fatty acids induce changes in immune responses by initiating various processes such as altering membrane fluidity, lipid peroxide formation, eicosanoid production, and gene regulation. With the combination of these processes and their effects on the immune system, dietary fatty acids influence pathogen clearance. In another study, omega-9 (ω-9) fatty acids have been proven to boost the high-density lipoprotein cholesterol while reducing low-density lipoprotein [
144]. Omega-9 fatty acids are categorized as polyunsaturated fatty acid (PUFA), which regulates prostaglandin synthesis and induces wound healing. This explains the relevance of ω-9 fatty acids being considered as a potential biomarker for vibriosis in brown-marbled groupers [
145], which is also supported by a study on
E. fuscoguttatus by Nurdalila et al. [
142] that reported an increased concentration of ω-9 compared to omega-6 (ω-6).
Confirmation of biomarker identity is the major bottleneck in metabolomics investigations. What can be deduced from the various discussion regarding the roles of metabolites that are found to be notably upregulated or downregulated in the hosts and the roles they play inactivation of an immune response is that these amino acids collectively provide a chance to enhance further the existing approaches used to combat fish disease. For example, the fish feed can be improved in various ways where the metabolites that strengthen the immunity of fish can be incorporated into meals. Apart from that, knowledge about compounds that seemingly inhibit the immune response is also crucial for researchers to find a way around the inhibition so that the natural immune response of fish is not interrupted. There are many ways to apply a multi-omics approach in the aquaculture industry to enhance fish health and improve the current technologies involved in curbing the fish disease.
Table 2. Compilation of studies on fish immune response using various omics approaches.
| Fish Species |
Pathogen |
Organ/Tissue Samples |
Method |
Reference |
| |
Transcriptomics |
| Crucian carp (Carassius auratus) |
Aeromonas hydrophila (bacteria) |
Head kidney |
Illumina Hiseq sequencer |
[146] |
Tilapia
(Oreochromis niloticus) |
Streptococcus iniae (bacteria) |
Spleen |
Illumina HiSeq 2000 instrument |
[100] |
Brown-marbled grouper
(Epinephelus fuscoguttatus) |
Vibrio vulnificus (bacteria) |
Gill and whole-body tissue |
Illumina HiSeq. 4000 |
[103] |
Turbot
(Scophthalmus maximus) |
Vibrio anguillarum (bacteria) |
Intestine |
Illumina HiSeq 4000 |
[147] |
| Orange-spotted grouper (Epinephelus coioides) |
Vibrio alginolyticus (bacteria) |
Whole body tissue |
Illumina HiSeq 2000 |
[102] |
| Rainbow trout (Oncorhynchus mykiss) |
Flavobacterium (bacteria) |
Spleen |
Illumina TruSeq |
[148] |
Soiny mullet
(Liza haematocheila) |
Streptococcus dysgalactiae (bacteria) |
Spleen |
Illumina HiSeq 2000 |
[149] |
| Rainbow trout (Oncorhynchus mykiss) |
Ichthyophthirius multifiliis (parasite) |
Gill |
Illumina HiSeq 2500 |
[99] |
Large yellow croaker
(Larimichthys crocea) |
Cryptocaryan irritans (parasite) |
Liver |
Illumina HiSeq2000 |
[114] |
| Orange-spotted grouper (Epinephelus coioides) |
Cryptocaryon irritans (parasite) |
Skin |
Illumina HiSeq 2500 |
[150] |
Striped snakehead
(Channa striata) |
Red-spotted grouper nervous necrosis virus (RGNNV) |
Striped snakehead fish cells (SSN-1) |
Illumina HiSeq 2000 |
[151] |
Atlantic salmon
(Salmo salar) |
Infectious salmon anemia virus (ISAV) |
Spleen |
Illumina MiSeq sequencer |
[152] |
Grass carp
(Ctenopharyngodon idellus) |
Grass carp reovirus (GCRV) |
Kidney |
Illumina NextSeq500 |
[98] |
Asian seabass
(Lates calcarifer) |
Nervous necrosis virus (NNV) |
Epithelial cells |
Illumina HiSeq™ 2000 |
[113] |
Koi
(Cyprinus carpio) |
Cyprinid herpesvirus 3 (CyHV3) |
Spleen |
Illumina HiSeq 2500 |
[153] |
Pacific cod
(Gadus microcephalus) |
General/Not specified |
Thymus and head kidney |
Illumina HiSeq 2000 platform |
[101] |
| |
Proteomics |
Brown-marbled grouper
(Epinephelus fuscoguttatus) |
Vibrio parahaemolyticus (bacteria) |
Blood |
2D gel electrophoresis,
Matrix-assisted laser desorption/ionization-time of flight-mass spectrometry (MALDI-TOF-MS/MS) analysis. |
[16] |
| Rainbow trout (Oncorhynchus mykiss) |
Yersinia ruckeri (bacteria) |
Intestine |
Micro Liquid chromatography coupled with electrospray ionization and quadrupole time of flight tandem- mass spectrometry (LC-ESI-qTOF-MS/MS) analysis. |
[154] |
Zebrafish
(Danio rerio) |
Aeromonas hydrophila (bacteria) |
Skin |
2D gel electrophoresis,
MALDI-TOF-MS analysis,
Liquid chromatography–mass spectrometry (LC-MS/MS) analysis. |
[125] |
| Yellow catfish (Pelteobagrus fulvidraco) |
Edwardsiella ictalurid (bacteria) |
Skin mucus |
LC-MS/MS analysis |
[155] |
Common carp
(Cyprinus Carpio) |
Aeromonas hydrophila (bacteria) |
Intestine |
LC-MS/MS analysis |
[119] |
Pufferfish
(Takifugu obscurus) |
Aeromonas hydrophila (bacteria) |
Spleen |
LC-MS/MS analysis |
[156] |
| Japanese flounder (Paralichthys olivaceus) |
Edwardsiella tarda (bacteria) |
Liver |
isobaric tags for relative and absolute quantification (iTRAQ) analysis,
LC-MS/MS analysis. |
[118] |
Atlantic salmon
(Salmo salar) |
Neoparamoeba
Perurans (parasite) |
Gill |
2D gel electrophoresis,
LC-MS/MS analysis |
[120] |
Lumpsucker
(Cyclopterus lumpus) |
General/Not specified |
Skin mucus |
2D gel electrophoresis,
LC-MS/MS analysis |
[157] |
Gilthead seabream
(Sparus aurata L.) |
General/Not specified |
Skin mucus |
2D gel electrophoresis,
Peptide mass fingerprinting-mass spectrometry (PMF-MS/MS) analysis,
LC-MS/MS analysis |
[158] |
| |
Metabolomics |
Atlantic salmon
(Salmo salar) |
Aeromonas salmonicida (bacteria) |
Kidney |
Nuclear magnetic resonance (H-NMR) analysis. |
[40] |
Brown-marbled grouper
(Epinephelus fuscoguttatus) |
Vibrio vulnificus (bacteria) |
Caudal fin |
Fourier-transform infrared spectroscopy (FTIR) analysis. |
[4] |
Zebrafish
(Danio rerio) |
Vibrio alginolyticus (bacteria) |
Whole body tissue |
Gas chromatography–mass spectrometry (GC-MS) analysis. |
[159] |
Brown marble grouper
(Epinephelus fuscoguttatus) |
Vibrio vulnificus (bacteria) |
Muscle tissue |
GC-MS analysis |
[136,142] |
Tilapia
(Oreochromis niloticus) |
Streptococcus iniae (bacteria) |
Liver |
GC-MS analysis |
[133] |
Tilapia
(Oreochromis niloticus) |
Streptococcus iniae (bacteria) |
Liver |
GC-MS analysis |
[160] |
Tilapia
(Oreochromis niloticus) |
Streptococcus agalactiae (bacteria) |
Liver |
ultraperformance liquid chromatography-tandem mass spectrometry (UPLC-MS) analysis. |
[161] |
Zebrafish
(Danio rerio) |
Edwardsiella tarda (bacteria) |
Muscle tissue |
GC-MS analysis |
[162] |
| Japanese puffer (Takifugu rubripes) |
Cryptocaryon irritans (parasite) |
Blood serum |
LC-MS analysis |
[163] |
Tiger puffer fish
(Takifugu rubripes) |
Myxosporea (parasite) |
Blood serum |
GC-MS analysis |
[164] |
| Mandarin fish (Siniperca chuatsi) |
Infectious spleen and kidney necrosis virus (ISKNV) |
Chinese perch brain cell line |
ultra-high-performance liquid chromatography-quadrupole time-of-flight mass spectrometry (UHPLC-Q-TOF/MS) analysis. |
[165] |
| Grouper |
Red-spotted grouper nervous necrosis virus (RGNNV) |
Spleen |
LC-MS analysis |
[166] |
| Crucian carp blood (Carassius auratus gibelio) |
Cyprinid herpesvirus 2 |
Blood |
LC-MS analysis |
[167] |