Proper functioning of the digestive system is ensured by coordinated action of the central and peripheral nervous systems (PNS). Peripheral innervation of the digestive system can be viewed as intrinsic and extrinsic. The intrinsic portion is mainly composed of the neurons and glia of the enteric nervous system (ENS). The extrinsic part is formed by sympathetic, parasympathetic, and sensory branches of the PNS with Schwann cells (SCs) being the chef glial cells. SCs are a crucial component of digestive tract innervation, and a great deal of research evidence highlights the important status of these glial cells in health and disease.
- Schwann cells
- digestive system
- pancreas
- cancer
1. Oral Cavity
2. Esophagus
3. Stomach
4. Pancreas
4.1. Pancreatic Innervation and Insights from 3D Imaging
4.2. Physiological Role of Innervation and SCs in Healthy Pancreas
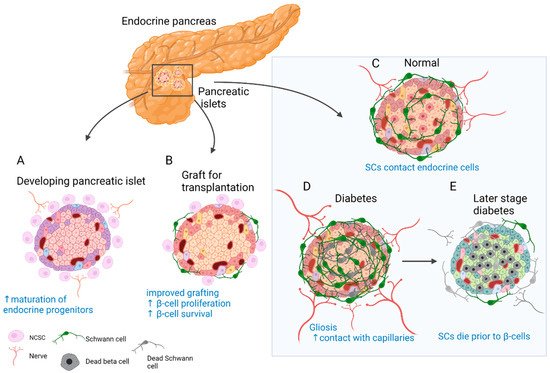
4.3. Fate and Function of SCs in Disorders of the Endocrine Pancreas
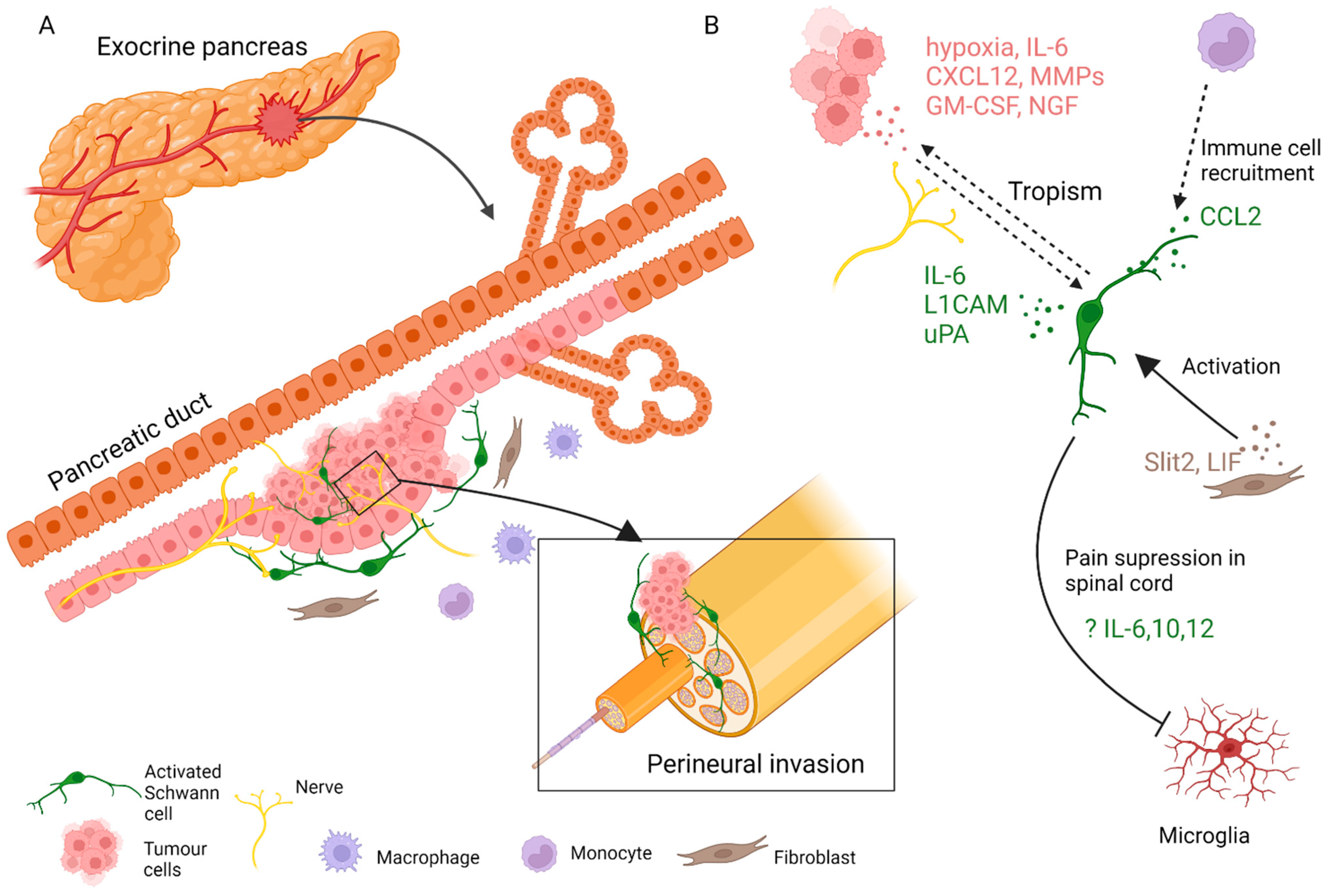
4.3. Fate and Function of SCs in Disorders of the Exocrine Pancreas
This entry is adapted from the peer-reviewed paper 10.3390/cells11050832
References
- Huff, T.; Daly, D.T. Neuroanatomy, Cranial Nerve 5 (Trigeminal); StatPearls Publishing: Treasure Island, FL, USA, 2021.
- Hargreaves, K.M. Orofacial pain. Pain 2011, 152, S25–S32.
- Rivera, C. Essentials of oral cancer. Int. J. Clin. Exp. Pathol. 2015, 8, 11884–11894.
- Viet, C.; Schmidt, B. Biologic Mechanisms of Oral Cancer Pain and Implications for Clinical Therapy. J. Dent. Res. 2011, 91, 447–453.
- Speight, P.M.; Farthing, P.M. The pathology of oral cancer. Br. Dent. J. 2018, 225, 841–847.
- Amit, M.; Takahashi, H.; Dragomir, M.P.; Lindemann, A.; Gleber-Netto, F.O.; Pickering, C.R.; Anfossi, S.; Osman, A.A.; Cai, Y.; Wang, R.; et al. Loss of p53 drives neuron reprogramming in head and neck cancer. Nature 2020, 578, 449–454.
- Zhang, Y.; Chen, M.; Liu, Z.; Wang, X.; Ji, T. The neuropeptide calcitonin gene-related peptide links perineural invasion with lymph node metastasis in oral squamous cell carcinoma. BMC Cancer 2021, 21, 1–10.
- Chiang, C.-Y.; Dostrovsky, J.O.; Iwata, K.; Sessle, B.J. Role of Glia in Orofacial Pain. Neuroscientist 2011, 17, 303–320.
- Ye, Y.; Salvo, E.; Romero-Reyes, M.; Akerman, S.; Shimizu, E.; Kobayashi, Y.; Michot, B.; Gibbs, J. Glia and Orofacial Pain: Progress and Future Directions. Int. J. Mol. Sci. 2021, 22, 5345.
- Salvo, E.; Saraithong, P.; Curtin, J.G.; Janal, M.N.; Ye, Y. Reciprocal interactions between cancer and Schwann cells contribute to oral cancer progression and pain. Heliyon 2019, 5, e01223.
- Salvo, E.; Tu, N.H.; Scheff, N.N.; Dubeykovskaya, Z.A.; Chavan, S.A.; Aouizerat, B.E.; Ye, Y. TNFα promotes oral cancer growth, pain, and Schwann cell activation. Sci. Rep. 2021, 11, 1840.
- Shan, C.; Wei, J.; Hou, R.; Wu, B.; Yang, Z.; Wang, L.; Lei, D.; Yang, X. Schwann cells promote EMT and the Schwann-like differentiation of salivary adenoid cystic carcinoma cells via the BDNF/TrkB axis. Oncol. Rep. 2015, 35, 427–435.
- Ein, L.; Mei, C.; Bracho, O.; Bas, E.; Monje, P.; Weed, D.; Sargi, Z.; Thomas, G.; Dinh, C. Modulation of BDNF–TRKB Interactions on Schwann Cell-induced Oral Squamous Cell Carcinoma Dispersion In Vitro. Anticancer Res. 2019, 39, 5933–5942.
- Kaukua, N.; Shahidi, M.K.; Konstantinidou, C.; Dyachuk, V.; Kaucka, M.; Furlan, A.; An, Z.; Wang, L.; Hultman, I.; Ährlund-Richter, L.; et al. Glial origin of mesenchymal stem cells in a tooth model system. Nature 2014, 513, 551–554.
- Martyn, G.V.; Shurin, G.V.; Keskinov, A.A.; Bunimovich, Y.L.; Shurin, M.R. Schwann cells shape the neuro-immune environs and control cancer progression. Cancer Immunol. Immunother. 2019, 68, 1819–1829.
- Couve, E.; Schmachtenberg, O. Schwann Cell Responses and Plasticity in Different Dental Pulp Scenarios. Front. Cell. Neurosci. 2018, 12, 299.
- Neuhuber, W.L.; Wörl, J. Enteric co-innervation of striated muscle in the esophagus: Still enigmatic? Histochem. Cell Biol. 2016, 146, 721–735.
- Kapitza, C.; Chunder, R.; Scheller, A.; Given, K.; Macklin, W.; Enders, M.; Kuerten, S.; Neuhuber, W.; Wörl, J. Murine Esophagus Expresses Glial-Derived Central Nervous System Antigens. Int. J. Mol. Sci. 2021, 22, 3233.
- Nikaki, K.; Sawada, A.; Ustaoglu, A.; Sifrim, D. Neuronal Control of Esophageal Peristalsis and Its Role in Esophageal Disease. Curr. Gastroenterol. Rep. 2019, 21, 59.
- Farmer, A.D.; Ruffle, J.K.; Aziz, Q. The Role of Esophageal Hypersensitivity in Functional Esophageal Disorders. J. Clin. Gastroenterol. 2017, 51, 91–99.
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer Incidence and Mortality Worldwide: Sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer 2015, 136, E359–E386.
- Pennathur, A.; Gibson, M.K.; Jobe, B.A.; Luketich, J.D. Oesophageal carcinoma. Lancet 2013, 381, 400–412.
- Griffin, N.; Rowe, C.W.; Gao, F.; Jobling, P.; Wills, V.; Walker, M.M.; Faulkner, S.; Hondermarck, H. Clinicopathological Significance of Nerves in Esophageal Cancer. Am. J. Pathol. 2020, 190, 1921–1930.
- Chen, J.-W.; Xie, J.-D.; Ling, Y.-H.; Li, P.; Yan, S.-M.; Xi, S.-Y.; Luo, R.-Z.; Yun, J.-P.; Xie, D.; Cai, M.-Y. The prognostic effect of perineural invasion in esophageal squamous cell carcinoma. BMC Cancer 2014, 14, 313.
- Xu, G.; Feng, F.; Liu, Z.; Liu, S.; Zheng, G.; Xiao, S.; Cai, L.; Yang, X.; Li, G.; Lian, X.; et al. Prognosis and Progression of ESCC Patients with Perineural Invasion. Sci. Rep. 2017, 7, 43828.
- Guo, Y.-N.; Tian, D.-P.; Gong, Q.-Y.; Huang, H.; Yang, P.; Chen, S.-B.; Billan, S.; He, J.-Y.; Huang, H.-H.; Xiong, P.; et al. Perineural Invasion is a Better Prognostic Indicator than Lymphovascular Invasion and a Potential Adjuvant Therapy Indicator for pN0M0 Esophageal Squamous Cell Carcinoma. Ann. Surg. Oncol. 2020, 27, 4371–4381.
- Tsunoda, S.; Okumura, T.; Ito, T.; Mori, Y.; Soma, T.; Watanabe, G.; Kaganoi, J.; Itami, A.; Sakai, Y.; Shimada, Y. Significance of nerve growth factor overexpression and its autocrine loop in oesophageal squamous cell carcinoma. Br. J. Cancer 2006, 95, 322–330.
- Zhou, Y.; Sinha, S.; Schwartz, J.L.; Adami, G.R. A subtype of oral, laryngeal, esophageal, and lung, squamous cell carcinoma with high levels of TrkB-T1 neurotrophin receptor mRNA. BMC Cancer 2019, 19, 1–12.
- Huang, S.-D.; Yuan, Y.; Liu, X.-H.; Gong, D.-J.; Bai, C.-G.; Wang, F.; Luo, J.-H.; Xu, Z.-Y. Self-renewal and chemotherapy resistance of p75NTR positive cells in esophageal squamous cell carcinomas. BMC Cancer 2009, 9, 9.
- Yamaguchi, T.; Okumura, T.; Hirano, K.; Watanabe, T.; Nagata, T.; Shimada, Y.; Tsukada, K. p75 neurotrophin receptor expression is a characteristic of the mitotically quiescent cancer stem cell population present in esophageal squamous cell carcinoma. Int. J. Oncol. 2016, 48, 1943–1954.
- Demir, I.E.; Boldis, A.; Pfitzinger, P.L.; Teller, S.; Brunner, E.; Klose, N.; Kehl, T.; Maak, M.; Lesina, M.; Laschinger, M.; et al. Investigation of Schwann Cells at Neoplastic Cell Sites Before the Onset of Cancer Invasion. J. Natl. Cancer Inst. 2014, 106, dju184.
- Espinosa-Medina, I.; Jevans, B.; Boismoreau, F.; Chettouh, Z.; Enomoto, H.; Müller, T.; Birchmeier, C.; Burns, A.J.; Brunet, J.-F. Dual origin of enteric neurons in vagal Schwann cell precursors and the sympathetic neural crest. Proc. Natl. Acad. Sci. USA 2017, 114, 11980–11985.
- Uesaka, T.; Nagashimada, M.; Enomoto, H. Neuronal Differentiation in Schwann Cell Lineage Underlies Postnatal Neurogenesis in the Enteric Nervous System. J. Neurosci. 2015, 35, 9879–9888.
- Chaudhry, S.R.; Liman, M.N.P.; Peterson, D.C. Anatomy, Abdomen and Pelvis, Stomach; StatPearls Publishing: Treasure Island, FL, USA, 2021.
- Phillips, R.J.; Powley, T.L. Innervation of the gastrointestinal tract: Patterns of aging. Auton. Neurosci. 2007, 136, 1–19.
- Deng, J.; You, Q.; Gao, Y.; Yu, Q.; Zhao, P.; Zheng, Y.; Fang, W.; Xu, N.; Teng, L. Prognostic Value of Perineural Invasion in Gastric Cancer: A Systematic Review and Meta-Analysis. PLoS ONE 2014, 9, e88907.
- Tao, Q.; Zhu, W.; Zhao, X.; Li, M.; Shu, Y.; Wang, D.; Li, X. Perineural Invasion and Postoperative Adjuvant Chemotherapy Efficacy in Patients with Gastric Cancer. Front. Oncol. 2020, 10, 530.
- Polli-Lopes, A.C.; Zucoloto, S.; Cunha, F.D.Q.; Figueiredo, L.A.D.S.; Garcia, S.B. Myenteric denervation reduces the incidence of gastric tumors in rats. Cancer Lett. 2003, 190, 45–50.
- Zhao, C.-M.; Hayakawa, Y.; Kodama, Y.; Muthupalani, S.; Westphalen, C.B.; Andersen, G.T.; Flatberg, A.; Johannessen, H.; Friedman, R.A.; Renz, B.W.; et al. Denervation suppresses gastric tumorigenesis. Sci. Transl. Med. 2014, 6, 250ra115.
- Hayakawa, Y.; Sakitani, K.; Konishi, M.; Asfaha, S.; Niikura, R.; Tomita, H.; Renz, B.W.; Tailor, Y.; Macchini, M.; Middelhoff, M.; et al. Nerve Growth Factor Promotes Gastric Tumorigenesis through Aberrant Cholinergic Signaling. Cancer Cell 2016, 31, 21–34.
- Langerhans, P.; Morrison, H. Contributions to the Microscopic Anatomy of the Pancreas; Bulletin of the Institute of the History of Medicine, The Johns Hopkins University Press: Hoboken, NJ, USA, 1937; Volume 5, pp. 259–297.
- Li, W.; Yu, G.; Liu, Y.; Sha, L. Intrapancreatic Ganglia and Neural Regulation of Pancreatic Endocrine Secretion. Front. Neurosci. 2019, 13, 21.
- Babic, T.; Travagli, R.A. Neural Control of the Pancreas; APA: Prairie Village, KS, USA, 2016.
- Makhmutova, M.; Caicedo, A. Optical Imaging of Pancreatic Innervation. Front. Endocrinol. 2021, 12, 445.
- Chien, H.-J.; Chiang, T.-C.; Peng, S.-J.; Chung, M.-H.; Chou, Y.-H.; Lee, C.-Y.; Jeng, Y.-M.; Tien, Y.-W.; Tang, S.-C. Human pancreatic afferent and efferent nerves: Mapping and 3-D illustration of exocrine, endocrine, and adipose innervation. Am. J. Physiol. Liver Physiol. 2019, 317, G694–G706.
- Alvarsson, A.; Jimenez-Gonzalez, M.; Li, R.; Rosselot, C.; Tzavaras, N.; Wu, Z.; Stewart, A.F.; Garcia-Ocaña, A.; Stanley, S.A. A 3D atlas of the dynamic and regional variation of pancreatic innervation in diabetes. Sci. Adv. 2020, 6, eaaz9124.
- Campbell-Thompson, M.; Tang, S.-C. Pancreas Optical Clearing and 3-D Microscopy in Health and Diabetes. Front. Endocrinol. 2021, 12, 644826.
- Ushiki, T.; Watanabe, S. Distribution and ultrastructure of the autonomic nerves in the mouse pancreas. Microsc. Res. Tech. 1997, 37, 399–406.
- Sunami, E.; Kanazawa, H.; Hashizume, H.; Takeda, M.; Hatakeyama, K.; Ushiki, T. Morphological Characteristics of Schwann Cells in the Islets of Langerhans of the Murine Pancreas. Arch. Histol. Cytol. 2001, 64, 191–201.
- Tsuchitani, M.; Sato, J.; Kokoshima, H. A comparison of the anatomical structure of the pancreas in experimental animals. J. Toxicol. Pathol. 2016, 29, 147–154.
- Rodriguez-Diaz, R.; Abdulreda, M.H.; Formoso, A.L.; Gans, I.; Ricordi, C.; Berggren, P.-O.; Caicedo, A. Innervation Patterns of Autonomic Axons in the Human Endocrine Pancreas. Cell Metab. 2011, 14, 45–54.
- Donev, S. Ultrastructural evidence for the presence of a glial sheath investing the islets of Langerhans in the pancreas of mammals. Cell Tissue Res. 1984, 237, 343–348.
- Love, J.A.; Yi, E.; Smith, T.G. Autonomic pathways regulating pancreatic exocrine secretion. Auton. Neurosci. 2007, 133, 19–34.
- Rodriguez-Diaz, R.; Caicedo, A. Neural control of the endocrine pancreas. Best Pract. Res. Clin. Endocrinol. Metab. 2014, 28, 745–756.
- Guyot, M.; Simon, T.; Ceppo, F.; Panzolini, C.; Guyon, A.; Lavergne, J.; Murris, E.; Daoudlarian, D.; Brusini, R.; Zarif, H.; et al. Pancreatic nerve electrostimulation inhibits recent-onset autoimmune diabetes. Nat. Biotechnol. 2019, 37, 1446–1451.
- Makhmutova, M.; Weitz, J.; Tamayo, A.; Pereira, E.; Boulina, M.; Almaça, J.; Rodriguez-Diaz, R.; Caicedo, A. Pancreatic β-Cells Communicate with Vagal Sensory Neurons. Gastroenterology 2021, 160, 875–888.e11.
- Lausier, J.; Diaz, W.C.; Roskens, V.; LaRock, K.; Herzer, K.; Fong, C.G.; Latour, M.G.; Peshavaria, M.; Jetton, T.L. Vagal control of pancreatic β-cell proliferation. Am. J. Physiol. Metab. 2010, 299, E786–E793.
- Borden, P.; Houtz, J.; Leach, S.D.; Kuruvilla, R. Sympathetic Innervation during Development Is Necessary for Pancreatic Islet Architecture and Functional Maturation. Cell Rep. 2013, 4, 287–301.
- Nekrep, N.; Wang, J.; Miyatsuka, T.; German, M.S. Signals from the neural crest regulate beta-cell mass in the pancreas. Development 2008, 135, 2151–2160.
- Plank, J.L.; Mundell, N.A.; Frist, A.Y.; LeGrone, A.W.; Kim, T.; Musser, M.A.; Walter, T.J.; Labosky, P.A. Influence and timing of arrival of murine neural crest on pancreatic beta cell development and maturation. Dev. Biol. 2011, 349, 321–330.
- Yang, Y.H.C.; Kawakami, K.; Stainier, D.Y. A new mode of pancreatic islet innervation revealed by live imaging in zebrafish. eLife 2018, 7, e34519.
- Persson-Sjögren, S.; Holmberg, D.; Forsgren, S. Remodeling of the innervation of pancreatic islets accompanies insulitis preceding onset of diabetes in the NOD mouse. J. Neuroimmunol. 2005, 158, 128–137.
- Taborsky, G.J., Jr.; Mei, Q.; Hackney, D.J.; Figlewicz, D.P.; Leboeuf, R.; Mundinger, T.O. Loss of islet sympathetic nerves and impairment of glucagon secretion in the NOD mouse: Relationship to invasive insulitis. Diabetologia 2009, 52, 2602–2611.
- Chiu, Y.-C.; Hua, T.-E.; Fu, Y.-Y.; Pasricha, P.J.; Tang, S.-C. 3-D imaging and illustration of the perfusive mouse islet sympathetic innervation and its remodelling in injury. Diabetologia 2012, 55, 3252–3261.
- Teitelman, G.; Guz, Y.; Ivkovic, S.; Ehrlich, M. Islet injury induces neurotrophin expression in pancreatic cells and reactive gliosis of peri-islet Schwann cells. J. Neurobiol. 1998, 34, 304–318.
- Tang, S.-C.; Chiu, Y.-C.; Hsu, C.-T.; Peng, S.-J.; Fu, Y.-Y. Plasticity of Schwann cells and pericytes in response to islet injury in mice. Diabetologia 2013, 56, 2424–2434.
- Tang, S.-C.; Peng, S.-J.; Chien, H.-J. Imaging of the islet neural network. Diabetes Obes. Metab. 2014, 16, 77–86.
- Winer, S.; Tsui, H.; Lau, A.; Song, A.; Li, X.; Cheung, R.K.; Sampson, A.; Afifiyan, F.; Elford, A.; Jackowski, G.; et al. Autoimmune islet destruction in spontaneous type 1 diabetes is not β-cell exclusive. Nat. Med. 2003, 9, 198–205.
- Olerud, J.; Kanaykina, N.; Vasilovska, S.; King, D.; Sandberg, M.; Jansson, L.; Kozlova, E.N. Neural crest stem cells increase beta cell proliferation and improve islet function in co-transplanted murine pancreatic islets. Diabetologia 2009, 52, 2594–2601.
- Grapensparr, L.; Vasylovska, S.; Li, Z.; Olerud, J.; Jansson, L.; Kozlova, E.; Carlsson, P.-O. Co-transplantation of Human Pancreatic Islets with Post-migratory Neural Crest Stem Cells Increases β-Cell Proliferation and Vascular and Neural Regrowth. J. Clin. Endocrinol. Metab. 2015, 100, E583–E590.
- Lau, J.; Vasylovska, S.; Kozlova, E.; Carlsson, P.-O. Surface Coating of Pancreatic Islets with Neural Crest Stem Cells Improves Engraftment and Function after Intraportal Transplantation. Cell Transplant. 2015, 24, 2263–2272.
- Juang, J.-H.; Kuo, C.-H.; Peng, S.-J.; Tang, S.-C. 3-D Imaging Reveals Participation of Donor Islet Schwann Cells and Pericytes in Islet Transplantation and Graft Neurovascular Regeneration. eBioMedicine 2015, 2, 109–119.
- Ngamjariyawat, A.; Turpaev, K.; Vasylovska, S.; Kozlova, E.N.; Welsh, N. Co-Culture of Neural Crest Stem Cells (NCSC) and Insulin Producing Beta-TC6 Cells Results in Cadherin Junctions and Protection against Cytokine-Induced Beta-Cell Death. PLoS ONE 2013, 8, e61828.
- Yantha, J.; Tsui, H.; Winer, S.; Song, A.; Wu, P.; Paltser, G.; Ellis, J.; Dosch, H.-M. Unexpected Acceleration of Type 1 Diabetes by Transgenic Expression of B7-H1 in NOD Mouse Peri-Islet Glia. Diabetes 2010, 59, 2588–2596.
- Daniel S. Longnecker; Anatomy and Histology of the Pancreas. The Pancreapedia: Exocrine Pancreas Knowledge Base 2014, NA, NA, 10.3998/panc.2014.3.
- Roberta De Angelis; Milena Sant; Michel Coleman; Silvia Francisci; Paolo Baili; Daniela Pierannunzio; Annalisa Trama; Otto Visser; Hermann Brenner; Eva Ardanaz; et al. Cancer survival in Europe 1999–2007 by country and age: results of EUROCARE-5—a population-based study. The Lancet Oncology 2014, 15, 23-34, 10.1016/s1470-2045(13)70546-1.
- Güralp Onur Ceyhan; Ihsan Ekin Demir; Ulrich Rauch; Frank Bergmann; Michael W Müller; Markus W Büchler; Helmut Friess; Karl-Herbert Schäfer; Pancreatic Neuropathy Results in “Neural Remodeling” and Altered Pancreatic Innervation in Chronic Pancreatitis and Pancreatic Cancer. American Journal of Gastroenterology 2009, 104, 2555-2565, 10.1038/ajg.2009.380.
- Stephan Schorn; Ihsan Ekin Demir; Bernhard Haller; Florian Scheufele; Carmen Mota Reyes; Elke Tieftrunk; Mine Sargut; Ruediger Goess; Helmut Friess; Güralp Onur Ceyhan; et al. The influence of neural invasion on survival and tumor recurrence in pancreatic ductal adenocarcinoma – A systematic review and meta-analysis. Surgical Oncology 2017, 26, 105-115, 10.1016/j.suronc.2017.01.007.
- Elisabeth Hessmann; Soeren M. Buchholz; Ihsan Ekin Demir; Shiv K. Singh; Thomas M. Gress; Volker Ellenrieder; Albrecht Neesse; Microenvironmental Determinants of Pancreatic Cancer. Physiological Reviews 2020, 100, 1707-1751, 10.1152/physrev.00042.2019.
- Dale E. Bockman; Markus Büchler; Hans G. Beger; Interaction of pancreatic ductal carcinoma with nerves leads to nerve damage. Gastroenterology 1994, 107, 219-230, 10.1016/0016-5085(94)90080-9.
- Ihsan Ekin Demir; Alexandra Boldis; Paulo L. Pfitzinger; Steffen Teller; Eva Brunner; Natascha Klose; Timo Kehl; Matthias Maak; Marina Lesina; Melanie Laschinger; et al. Investigation of Schwann Cells at Neoplastic Cell Sites Before the Onset of Cancer Invasion. JNCI: Journal of the National Cancer Institute 2014, 106, NA, 10.1093/jnci/dju184.
- Ihsan Ekin Demir; Elke Tieftrunk; Stephan Schorn; Ömer Cemil Saricaoglu; Paulo L Pfitzinger; Steffen Teller; Kun Wang; Christine Waldbaur; Magdalena U Kurkowski; Sonja Maria Wörmann; et al. Activated Schwann cells in pancreatic cancer are linked to analgesia via suppression of spinal astroglia and microglia. Gut 2016, 65, 1001-1014, 10.1136/gutjnl-2015-309784.
- Ihsan Ekin Demir; Kristina Kujundzic; Paulo L. Pfitzinger; Ömer Cemil Saricaoglu; Steffen Teller; Timo Kehl; Carmen Mota Reyes; Linda S. Ertl; Zhenhua Miao; Thomas J. Schall; et al. Early pancreatic cancer lesions suppress pain through CXCL12-mediated chemoattraction of Schwann cells. Proceedings of the National Academy of Sciences 2016, 114, 201606909-E94, 10.1073/pnas.1606909114.
- Bakst, R.L.; Xiong, H.; Chen, C.-H.; Deborde, S.; Lyubchik, A.; Zhou, Y.; He, S.; McNamara, W.; Lee, S.-Y.; Olson, O.; et al. Inflammatory Monocytes Promote Perineural Invasion via CCL2-Mediated Recruitment and Cathepsin B Expression. Cancer Res. 2017, 77, 6400–6414.
- Secq, V.; Leca, J.F.; Bressy, C.; Guillaumond, F.; Skrobuk, P.; Nigri, J.; Lac, S.; Lavaut, M.-N.; Bui, T.-T.; Thakur, A.K.; et al. Stromal SLIT2 impacts on pancreatic cancer-associated neural remodeling. Cell Death Dis. 2015, 6, e1592.
- Bressy, C.; Lac, S.; Nigri, J.; Leca, J.; Roques, J.; Lavaut, M.-N.; Secq, V.; Guillaumond, F.; Bui, T.-T.; Pietrasz, D.; et al. LIF Drives Neural Remodeling in Pancreatic Cancer and Offers a New Candidate Biomarker. Cancer Res. 2017, 78, 909–921.