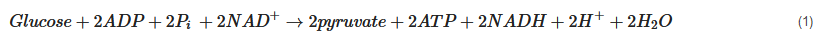Traumatic brain injury (TBI) is a health burden affecting over 50 million people each year globally [
1,
2]. Its long-term adverse effects to the brain may create a significant burden implicating social integration, independence, and employability. Population-based studies have reported that 238,000 people each year sustain a TBI in the UK, and it is the overall leading cause of death in young adults (25–35 years) in developed countries [
3]. In 2013, TBIs in the USA led to about 2.5 million emergency department visits, 282,000 hospitalisations, and 56,000 deaths [
4]. Mild or severe TBI patients are at risk of secondary brain injury that arises from complex physiological responses to the initial injury. These complex physiological processes are thought to include a series of sequential events including altered cerebral metabolism. Monitoring cerebral metabolism along with other protocols to identify early changes and prevent secondary brain injuries during neurocritical care management can improve outcomes of severe TBI patients [
5,
6]. Currently, cerebral metabolism is monitored using cerebral microdialysis, an invasive technique in which interstitial brain fluid can be sampled. The sampled fluid (also termed microdialysate) is usually analysed hourly offline using a bedside analyser, for which its operating principle is based on enzymatic reactions [
7,
8]. However, the data analysed are retrospective, hindering real-time interventions. Thus, effective and timely measurements of the microdialysates are required to predict the onset of secondary brain damage. To fulfil this, emerging research has integrated microdialysis with biosensors for continuous online detection of relevant cerebral metabolites found in microdialysates. The intent of these technologies is to provide accurate and actionable information that may help inform clinical decision making and aid in the prognostication of patients’ outcome.
2. Importance of Monitoring Brain Metabolism for TBI
2.1. Traumatic Brain Injury (TBI)
TBI is an insult to the brain due to an external force causing damage to the brain’s structure and function [
9]. It represents mild, moderate, and severe effects of physical assault to the brain that may cause sequential, primary, or secondary ramifications. Thus, TBI can be classed into a primary mechanical injury, which is non-amenable to medical treatment and a delayed secondary brain injury involving various changes at the cellular and molecular level that could in theory be counteracted. The sudden mechanical injury is a widespread tearing, shearing, or stretching of axons, referred to as axonal injury, as well as contusions, hemorrhages, and lacerations [
10]. Delayed secondary injury evolves hours or days after the initial primary mechanical trauma due to neuronal and glial dysfunction, neuroinflammation, cerebral oedema, and metabolic changes. Consequently, this leads to various physiologic alterations including hypoperfusion, blood–brain barrier (BBB) disruption, oxidative injury, and mitochondrial dysfunction [
11,
12,
13,
14,
15,
16]. This complexity of secondary brain injury poses diagnostic and therapeutic challenges; therefore, additional invasive monitoring interventions are required during neurocritical care of severe TBI patients. These additional metrics may provide insights into brain tissue oxygenation, intracranial pressure (ICP), cerebral perfusion pressure (CPP), electrophysiology, and local brain metabolism [
11]. Abnormal local brain metabolism is linked to poor patient clinical outcomes, thus interrogating the brain to monitor its chemistry is necessary [
17].
2.2. Cerebral Metabolism
Evidence shows that cerebral metabolism is disturbed following TBI, although the exact mechanisms are incompletely understood due to the complex and heterogeneous nature of TBI [
7]. The brain uses glucose as a preferred substrate for energy consumption, so the regulation of cerebral glucose metabolism is crucial. Oxidative metabolism of glucose provides most of the ATPs utilised by the brain. However, biosynthetic routes that branch from the glycolytic pathway and the tricarboxylic acid (TCA) cycle and other pathways including the pentose phosphate shunt, glucose storage as glycogen, and the malate-aspartate shuttle all have significant roles [
18]. Understanding of the altered cerebral metabolism is incomplete without knowledge of the glucose metabolism in an uninjured brain.
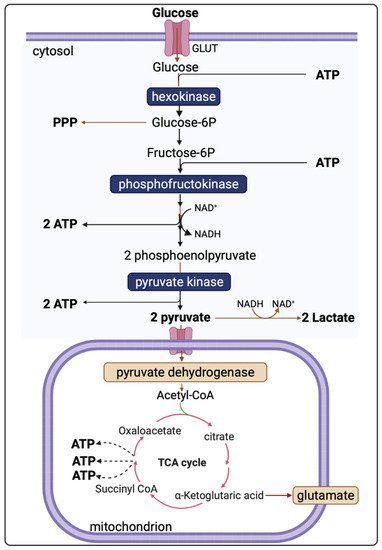
Figure 1 presents the major energy pathways in the brain.
In the uninjured brain, glucose uptake by both neuronal and glial cells undergoes oxidation via glycolysis to form two pyruvate molecules. Glycolysis is independent of oxygen and, thus, can occur under either aerobic or anaerobic conditions. The energy yield of ATP per molecule of glucose depends on whether mitochondrial shuttle mechanisms are operational or not. To generate one molecule of pyruvate by glycolysis, one molecule of NAD
+ is converted into NADH, which must be recycled (oxidised) back to NAD
+ if glycolysis is to be sustained. One recycling mechanism is by the electron-transport chains of mitochondria (if operational). As NADH cannot cross the mitochondrial membrane, the requisite hydrogens and electrons are transferred indirectly by “shuttle” mechanisms-malate-aspartate shuttle system and/or the glycerol-3-phosphate shuttle [
19]. NADH can also be recycled to NAD
+ by the lactate dehydrogenase (LDH)-mediated conversion of pyruvate to lactate in the cytosol, as an epilogue to “anaerobic” glycolysis.
The overall reaction of glycolysis and net yield of ATP molecules is presented in Equation (1).
Pyruvate, the end-product of glycolysis, can enter mitochondria where it is converted to acetyl-CoA by pyruvate dehydrogenase. Acetyl-CoA is further metabolised in the tricarboxylic acid (TCA) cycle in mitochondria. The sum of all the reactions in the TCA cycle is presented in Equation (2).
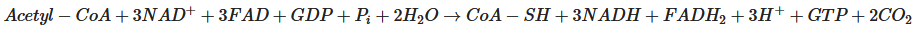
Subsequently, NADH and FADH
2 are utilised by the mitochondrial electron transport chain (by Complexes I and II, respectively); electrons are transferred to complexes III and IV, where O
2 is the terminal electron acceptor on Complex IV, followed by ATP synthesis by ATP synthase (also termed Complex V) [
20]. The yield per molecule of glucose metabolised fully to CO
2 (by combined glycolysis, NADH shuttling, and mitochondrial respiration) is theoretically 36–38 ATP molecules. However, the actual yield is considered somewhat lower [
21,
22].
2.3. Altered Cerebral Metabolism Due to TBI
Imbalance in cerebral glucose metabolism following TBI is well-documented [
7,
23,
24] and attributable, at least partly, to the altered ATP production in the brain’s major energy pathways. Due to a high energy demand following TBI, abnormally low levels (<0.8 mM) of extracellular glucose occur [
17], possibly because of upregulated glucose uptake by neurones and glia. Conversely, neurones and glia may sometimes be too damaged to take up glucose from extracellular fluid, leading to hypometabolism characterised by abnormally high extracellular glucose. Thus, there is an optimum extracellular glucose range, although there is insufficient evidence to define this exactly [
25]. Extracellular lactate can also be utilised as an alternative fuel [
24].
13C-labelled microdialysis studies have demonstrated that the traumatically injured brain uses lactate via the TCA cycle [
24,
26]. Another
13C-labelled microdialysis study found lactate production from 1,2-
13C
2 glucose via glycolysis and to a lesser extent via PPP [
23]. Lactate was also identified as a spin-out product (cataplerosis) from the TCA cycle in
13C-labelled microdialysis studies using 2,3-
13C
2 succinate as a substrate [
11,
23].
A persistent high lactate/pyruvate ratio (LPR) (LPR > 25 or >40) indicates metabolic dysfunction or crisis. In a microdialysis study of 233 TBI patients, acute-phase LPR > 25 was associated with poor clinical outcomes 6 months later [
17]. High LPR, despite seemingly adequate oxygen and glucose delivery to brain tissues, is regarded as indicating mitochondrial dysfunction [
11,
27]. The concentrations of lactate and pyruvate and their ratio (LPR) provide useful information about the cellular redox state in the region of interest. The extracellular LPR is thought to reflect the LPR in the cytoplasm—itself in equilibrium with cytoplasmic NADH/NAD
+ ratio [
28]. Glucose, lactate, pyruvate, and LPR were cited as the most clinically relevant biomarkers in a consensus statement from the 2014 International Microdialysis Forum [
25]. Timely assessment of these is, therefore, essential in the early detection of secondary brain injury allowing prompt interventions.
Table 1 summarises neuroprotective interventions for altered neurochemistry.
3. Review of Sensor Technologies for Brain Metabolism
In selecting scholarly articles to discuss here, emphasis was placed on work published in the areas of biosensors, TBI, cerebral metabolism, and microdialysis in the last 10 years. Table 2 summarises some of the most relevant articles discussed here. Those biosensors are in the research and development phases and are not approved for routine clinical use.