During the early phases of the development of the adaptive immune system, only T lymphocytes, which are capable of recognizing non-self-peptides are selected. This negative selection allows the activation of the effector functions of surveying T lymphocytes, only when their T cell receptor (TCR) specifically interacts with a foreign peptide presented by a major histocompatibility complex (MHC) protein on an antigen-presenting cell (APC).
2. Antigen Processing and Presentation in Teleost Fish
The demonstration of APP occurrence in teleost fish came from studies with the channel catfish (
Ictalurus punctatus) with evidence indicating that MHC-like molecules govern APP with a similar restriction as observed in mammals [
32] and with exogenous antigens being endocytosed by APC and inducing the proliferation of lymphocytes [
33]. There is sufficient evidence supporting that the functions of MHC class I and II molecules in fish and mammals are similar [
34]. A scheme of mammal APP indicating the key molecular players that have been studied and characterized in teleost fish is shown in
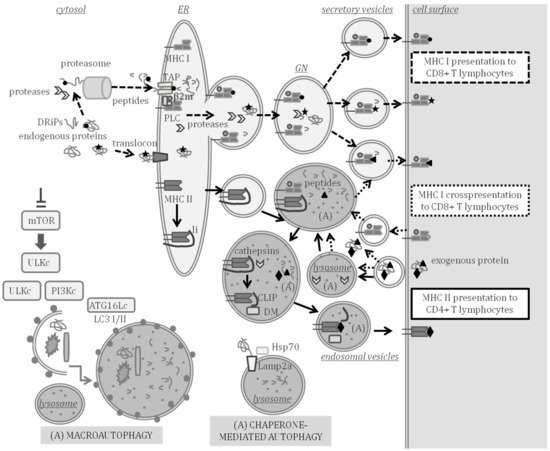
Figure 1, indicating the compartments where APP and autophagy can take place at the same time.
Figure 1. Major histocompatibility complex (MHC) class I and II antigen processing and presentation (APP) and autophagy. MHC class I presents endogenous peptides produced in the cytosol (peptide represented by a circle) or in secretory compartments (star) to CD8 + T lymphocytes through the pathways indicated with the dashed arrows. Endogenously synthesized proteins and defective ribosomal products (DRiPs) are processed to peptides in the cytosol by the proteasome and cytosolic proteases. Peptides are then transported by transporter associated to antigen processing (TAP) to the ER where the peptide loading complex (PLC) is formed by a peptide, TAP, MHC class I, β2-microglobulin (β2m) and other assistant proteins. Endogenous proteins can enter the lumen of the ER through the translocon and be processed by proteases of the ER or the Golgi network (GN), thus, producing peptides that can also be loaded on MHC class I beyond the ER. Loaded MHC class I molecules are transported to the cell surface in secretory vesicles. MHC class I can also cross present peptides and prime CD8+ T lymphocytes through the pathway indicated with the dotted arrows. Cross-presented peptides (triangle) are produced by processing exogenous antigens in the cytosol or in secretory or endocytic compartments in which autophagy (A) may take place. MHC class II presents peptides (diamond) processed from exogenous proteins to CD4 + T lymphocytes through the pathway indicated with the continuous lined arrows. In the ER, MHC class II molecules are assembled and bind to the invariant chain li. The MHC class II is exported to endosomal compartments, in which autophagy can take place and where class II-associated invariant chain peptide (CLIP) remains bound to MHC class II after cleavage of li by lysosomal cathepsins. HLA-DM assists in the exchange of CLIP by a peptide that is exported to the cell surface through exocytic vesicles. Chaperone-mediated autophagy allows the targeting of cytosolic proteins to be degraded in lysosomes and is assisted by Lamp2a, Hsp70 and other chaperones. Macroautophagy is the engulfment of diverse cytosolic material to be degraded, induced by inhibition of the mammalian target of rapamycin (mTOR). Several multiprotein complexes (ULK, PI3K and ATG16L1) are involved in the formation of vesicles and lipidation of LC3-I to LC3-II, thereby, forming autophagosomes that fuse with lysosomes.
2.1. Antigen Processing and Presentation Genes Conserved in Teleost Fish
The emergence of adaptive immunity is related to the highly polymorphic
mhc genes encoding MHC, which in most jawed vertebrates are linked, scattered throughout a chromosomal genomic region with varying levels of recombination that also encodes other genes related to APP. Teleost fish are, however, the exception, as the
mhc class I gene family is expanded and located on several chromosomes, and the
mhc class II genes are lost in some fish species [
35,
36]. In spite of the loss of key components of the adaptive machinery, teleost fish are capable of mounting a robust immune response to pathogens [
5].
Interestingly, the polymorphism in
mhc class I observed in many teleosts is proposed to be driven by additional evolutionary pressures apart from pathogen resistance [
34]. Initial studies in comparative genomics with medaka (
Oryzias latipes) revealed the evolutionarily conserved core in teleost APP is
mhc class I and its related genes [
37] with the genes encoding MHC class I, proteasome subunits LMP2, LMP7 and TAP2 appearing as linked in linkage analysis [
37]. Genes related to MHC class I APP that have been cloned and characterized in teleost fish include several proteasome subunits [
38,
39,
40],
β2m [
41] or the ERp57 gene
pdia3 [
42,
43] and the tapasin gene
tapbp [
44].
The
mhc class I genes are organized in teleost fish in
U,
Z,
L,
S or
P lineages, although not all species have all lineages. Only the
U lineage is predicted to encode the genes that provide APP functions, displaying haplotypic variation and sharing conserved synteny with the core
MHC that is observed in mammalian genomes [
45]. The
mhc class I genes have been cloned and characterized in a wide number of teleost fish [
41,
46,
47,
48]. The zebrafish (
Danio rerio) shows substantial diversity within genes of proteasome subunits,
tap and
mhc class I genes [
49], indicating teleosts have polymorphisms within the MHC class I APP steps of cleavage, transport and presentation, all predicted to alter peptide specificity.
Vertebrate genome duplication events in evolution have provided cyprinids, salmonids and neoteleosts with duplicates of genes involved in MHC class I APP and therefore with the potential of different PLC [
50]. The
U lineage in the zebrafish is located on chromosome 19, although two
U genes map to chromosome 22 and are predicted to play non classical roles in APP because they lack key residues for peptide binding and display limited polymorphic variation [
51]. Further studies are needed to decipher the complexity in the number of functional genes involved in MHC class I APP, as gene duplicates respond differently to stimulation and may not share biological functions [
52].
The
mhc class II genes in teleost fish are classified in three major groups (
DA,
DB and
DE), with only
DA exhibiting the classical features [
45]. The gene coding for the α chain of class II (
spau-daa) has been characterized in the gilthead seabream (
Sparus aurata), showing conserved important features and a high variability in the α1 domain that forms the peptide-binding domain (PBD) [
53]. The
mhc class II loci have been lost independently in gadiform fishes, such as Atlantic cod (
Gadus morhua) [
54] and in some syngnathiform fishes, such as the pipefish (
Syngnathus typhle) [
55].
HLA-DM, which assists MHC class II in peptide loading, is highly conserved in tetrapod species. The
HLA-DM-lineage had only been reported to the level of amphibians until it was recently reported to exist in lungfish [
56]. A critical Trp residue required for the HLA-DM system found in mammals is not encoded in classical teleost
mhc class II and only found in non-classical
mhc class II genes [
57]. The invariant chain (li) and cathepsins, which are involved in the cleavage of li have been studied in depth [
58,
59,
60,
61,
62,
63,
64,
65,
66]. Conserved motifs in the CLIP region of li exist throughout jawed vertebrates. Conserved li residues and cathepsin orthologs suggest their long co-evolution in the APP, with sub-functional duplicated
cd74 genes [
56,
67]. In rainbow trout (
Oncorhynchus. mykiss), two
cd74 genes have been characterized, and their expression is regulated differently [
58,
59].
2.2. Antigen-Presenting Cells (APC) and Professional APC (pAPC) in Teleost Fish
All cells are APC as they express MHC class I and can perform APP. Indeed, MHC class I molecules was reported to be expressed in teleost fish in similar cell types as in higher vertebrates [
68], and there is evidence indicating that the activation of CD8+ T lymphocyte-mediated cytotoxicity is MHC class I restricted [
34,
69]. MHC class II is, however, only expressed in the professional APC (pAPC), which are in charge of the initial activation or priming of effector T lymphocytes.
These pAPC mainly reside in lymphoid organs, such as the head-kidney, the thymus or the spleen, and are capable of priming T lymphocytes to become cytotoxic CD8+ T lymphocytes or CD4+ T helper lymphocytes. The role of pAPC is thus to act as immune sentries that engulf pathogens and carry foreign antigens to lymphoid tissues in order to stimulate an antigen-specific T lymphocyte response. As described in higher vertebrates, the pAPC in some teleost fish species are dendritic cells (DCs), B cells and macrophages, all of which have been characterized in depth in the zebrafish [
70]. However, in other species of bony fishes, in addition to DCs, B cells and macrophages, granulocytes [
53] and erythrocytes (red blood cells, RBC) [
71,
72,
73] also have a prominent role in APP.
The phenotypic and functional features of DCs have been studied in several teleost species, and methods to enrich DCs have been established. In zebrafish, a pAPC subset strongly resembling mammalian DCs was characterized in relation to affinity for the lectin peanut agglutinin (PNA) [
74]. Myeloid cells that were PNA(hi) had phagocytic activity and expressed genes associated to DC function and APP, including MHC class II [
74]. Most importantly PNA(hi) cells could activate T lymphocytes in an antigen-dependent manner [
74].
Among the phenotypic cell surface markers of zebrafish DCs are MHC class II, CD80/86, CD83 and CD209, and all these molecules were involved in the activation of CD4 + T lymphocytes [
75]. In barramundi (
Lates calcarifer), maturation of spleen- and pronephros-derived monocytes exposed in vitro to bacterial-derived peptidoglycans induced differentiation to DC. These DC were phagocytic and could migrate towards pathogens or pathogen associated patterns, they expressed MHC class II and DC-SCRIPT, a conserved zinc finger protein preferentially expressed in human DC subtypes [
76]. Importantly, barramundi DCs induced proliferation of effector T lymphocytes [
76].
In rainbow trout, mucosal tissues from skin and gills have a DC subpopulation that co-expressed CD8α and MHC class II [
77], with a homologous population described in intestinal lamina propia [
78]. However, the intestinal CD8+ DCs exhibited phenotypic and functional differences compared to skin and gill CD8+ DCs, suggesting the presence of different subpopulations of DCs on different locations [
78]. Furthermore, two phenotypes of pAPC were characterized in the gills of rainbow trout. One had features of a monocyte/macrophage/dendritic cell-type; expressing CD45, CD83 and IL-1 β, while the second phenotype expressed almost all genes related to the phagosome, lysosome and APP pathways and exhibited similar characteristics to mammalian M cells, which play a role in mucosal immune surveillance [
79].
In a different species, the orange-spotted grouper (
Epinephelus coioides), a transcriptomic study performed on skin after
Cryptocaryon irritans infection, revealed upregulation of DCs markers, including CD209 and CD83 [
80]. CD207 is a marker of Langerhans cells (tissue-resident macrophages of the skin), which are developmentally similar to DCs. Infection of the grass carp (
Ctenopharyngodon idella) with
Flavobacterium columnare upregulated CD207 only in the spleen, suggesting that the biological function of CD207 may be related to APP [
81].
B cells are pAPC in teleosts as they can uptake particulate antigens by phagocytosis, process and present antigens to T lymphocytes [
82,
83]. In bony fish, B cells produce three isotypes of immunoglobulins (Ig): IgM, IgD and IgT/Z [
82]. In accordance, three main B-cell lineages have been described in teleosts, a B-cell lineage expressing IgT/Z, a lineage expressing IgD and the most common B-cell lineage co-expressing IgD and IgM [
82].
In the rainbow trout, infection in vitro of blood or spleen IgM+ B cells with viral hemorrhagic septicemia virus (VHSV) upregulates MHC class II, CD80/86 or CD83, suggesting the activation towards a pAPC profile [
84]. In vivo IgM+ B cells are recruited to the peritoneum cavity after challenge with virus VHSV or bacteria
Escherichia coli and differentiate to cells specialized in APP [
85]. A teleost B cell subset has been proposed to be a counterpart of mammalian B1-B cells, which link innate and adaptive immunity [
86].
Professional phagocytic cells have a role in pAPC in teleost fish. Thus, macrophages are pAPC that participate in immunity as phagocytic cells, with two polarization states, the pro-inflammatory M1-type and anti-inflammatory M2-type. The expression of genes involved in APP was detected in both M1- and M2-type macrophages in the zebrafish [
87]. In addition, the infection of macrophages upregulated MHC class II and transcripts of other genes involved in APP [
87]. In addition to macrophages, head-kidney granulocytes can express high levels of MHC class II in gilthead seabream [
53] or in Atlantic salmon (
Salmo salar) [
71] and are proposed as a model to study autophagy and APP during vertebrate immune response [
71].
2.3. Peptide Processing, Loading and Transport by MHC in Teleost Fish
A number of studies have characterized some of the key proteins involved in the processing of antigens to peptides, the loading on MHC and MHC/peptide transport to the cell surface of APC. Concerning the MHC class I APP pathway, in European sea bass (
Dicentrarchus labrax), different transcripts of MHC class I have been identified per individual, detecting variability at the PBD, with a three-dimensional homology model that is consistent with that of other vertebrates [
41]. The proteasome activator subunits PA28α and PA28β were upregulated upon immune stimulation in the Japanese flounder (
Paralichthys olivaceus) [
38], the large yellow croaker (
Larimichthys crocea) [
39] and the rock bream (
Oplegnathus fasciatus) [
40].
The expression of β2m was induced by an antigen in the European sea bass [
41]. The interaction of tapasin with TAP, MHC class I and ERp57, all which are known to occur during assembly of the mammalian peptide loading complex (PLC), was conserved in the monocyte/macrophage rainbow trout cell line RTS11 [
88]. Homology models of the protein ERp57 in European sea bass support that they are orthologs of the mammalian protein involved in MHC class I APP [
42].
In rainbow trout, expression analysis supports the role of ERp57 during the activation of the immune response in teleosts [
43], although some features conserved in mammalian ERp57 are not conserved in the fish gene [
43]. In the same species, tapasin was mainly expressed in immune system organs and in cell lines derived from epithelial cells from the gill, liver and intestine [
44]. In addition, tapasin was strongly expressed in parallel with MHC class I in the RTS11 monocyte/macrophage derived cell line after viral infection [
44].
Concerning the MHC class II APP pathway, the expression of the alpha chain of gilthead seabream MHC class II (
Spau-DAA) was observed in peritoneal exudate leucocytes, head-kidney, spleen, thymus and gill; and the incubation of head-kidney leucocytes with yeast or bacteria upregulated levels of
Spau-DAA, whereas concanavalin A (ConA), lipopolysaccharide (LPS) or phytohemagglutinin A (PHA) did not [
53].
In the MHC class II APP, the interferon-γ-inducible lysosomal thiol reductase (GILT) is important as it reduces disulfide bonds in the unfolding of proteins and facilitates their subsequent cleavage by proteases. In zebrafish, purified recombinant GILT was capable of catalyzing the reduction of protein disulfide bonds [
89]. GILT was constitutively expressed in several tissues of zebrafish, large yellow croaker and mandarin fish (
Siniperca chuatsi) and upregulated in the spleen and kidney after stimulation [
89,
90,
91].
The invariant chain li, a key player in the formation of mammalian MHC class II/peptide complex, appears to be expressed independently of immune function in some teleost fish. The invariant chain-like proteins S25-7 and INVX of the rainbow trout were not expressed in the head-kidney, and their expression in spleen and gill was unaffected by immune stimulation [
59]. Cathepsins, the family of lysosomal cysteine proteases that are involved in the processing of li, have been characterized in depth in teleost species.
Several cathepsins have been characterized and found to be widely expressed and produced in vitro to confirm their protease activity: cathepsins D and L in the half-smooth tongue sole (
Cynoglossus semilaevis) [
61]; in the large yellow croaker cathepsin L and S [
62] and B [
60]; in the miiuy croaker (
Miichthys miiuy) cathepsin S [
63] and cathepsin B and cathepsin H [
64] were assessed; while cathepsin B was studied in the golden pompano (
Trachinotus ovatus) [
66] and cathepsin S in the channel catfish [
65]. The capacity of cathepsins to cleave the invariant chain li was studied in the large yellow croaker, finding that cathepsin B efficiently cleaves recombinant li in a time-dependent manner [
60].
Cathepsin S and cathepsin L both had cysteine protease activity and could remove propeptides and release active mature peptides; however, li inhibited the autocatalytic activation of cathepsin L, whereas activated cathepsin S could efficiently process li in vitro [
62]. These results suggest that cathepsins S and B are the main cathepsins involved in the processing of li in teleost fish [
60,
62]. It should be noted that, in the channel catfish, two cathepsin S genes have been characterized, (
ctss and
ctssa), both upregulated during bacterial infection, however, with differences between their expression profiles in mucosal surfaces, suggesting that they may exert disparate roles in mucosal immunity [
65].
3. Autophagy and Related LC3-Associated Phagocytosis in Teleost Fish
Autophagy is a self-digestion process that is highly conserved in evolution and involves lysosomal degradation of cytoplasmic material [
1]. Autophagy mainly occurs by fusion of autophagosomes, which engulf portions of cytosol and fuse with lysosomes (macroautophagy), or assisted by chaperones (chaperone-mediated autophagy, CMA) that deliver cytosolic proteins to lysosomes. One of the non-canonical functions of autophagy is to participate in APP for MHC class I and II activation of T lymphocytes [
3]. LC3-associated phagocytosis (LAP), however, is not a form of autophagy; it depends on key autophagy molecular players [
30].
3.1. Chaperone-Mediated Autophagy in Teleost Fish
CMA was long believed to only exist in birds and mammals [
92]; however, it has been proved to exist also in fish [
93,
94]. Lamp2a is the product of one of the three splice variants (
lamp2a,
lamp2b and
lamp2c) of gene
lamp2 that is transcribed by alternative splicing, and this genomic organization has been highly conserved during vertebrate evolution [
93]. The gene
lamp2 appears at the root of the vertebrate lineage with a Ct domain GYXXF sequence that is conserved in most teleosts [
93], including medaka. However,
lamp2 is divergent in zebrafish, which raised the question on the ability of this species to perform CMA [
92].
In medaka in vitro studies, the splice variant
lamp2a was found to control lysosomal accumulation of a reporter of CMA in a fibroblast cell line; and in vivo, a
lamp2a medaka knock-out exhibited carbohydrate and fat metabolism alterations [
93]. In cultured cells of the yellowtail fish (
Seriola quinqueradiata), three cytosolic members of the Hsp70 family have been isolated; heat-shock cognate proteins (Hsc)70-1 and Hsc70-2 and heat shock protein (Hsp)70 [
95]. Heat-induced expression of Hsp70, together with the lysosome localization of Hsc70/Hsp70 suggests that CMA is induced by heat shock in bony fish [
95]. In the zebrafish, a member of the small heat shock protein (sHSP) family, Hspb8, interacts with proteins Bag3 and Hsc70, which are essential for the formation of an autophagy-inducing complex [
96].
3.2. Macroautophagy in Teleost Fish
Xenophagy is the capture and destruction of invading pathogens by means of a form of selective macroautophagy that requires the participation of receptors Sqstm1/p62 that bind to ubiquitinated cargo and LC3-II to form p62-droplets that accumulate at the surface of autophagosomes during their formation [
1,
30,
31]. The contribution of zebrafish models to study the specific elimination of certain intracellular substrates by autophagic pathways, has been recently reviewed [
97], as well as the role of selective autophagy in immunity to bacterial pathogens (
Shigella flexneri and
Mycobacterium marinum) [
98].
Inflammatory toxicity in zebrafish induces the cleavage of p62 by caspase-6 at a highly conserved D256 cleavage site, indicating certain stress stimuli can modulate autophagy through the caspase 6-p62 axis [
99]. Zebrafish mutant lines have shown that
becn1 is essential for autophagy, as knock-out mutants for this gene die at the larval stage [
100]. In addition, mutants in
becn1 and
atg7 have outlined the importance of
becn1 for maintaining homeostasis of nutrient metabolism and in liver development [
101].
3.3. LC3-Associated Phagocytosis in Teleost Fish
In a zebrafish embryo model, the interaction of human pathogen
Salmonella typhimurium with the autophagy machinery of macrophages identified LAP as the major host protective autophagy-related pathway responsible for macrophage defense during systemic infection [
102]. In contrast, in an infection model to study the human pathogen
Staphylococcus aureus, LAP was found to be involved in the intracellular handling of the pathogen by neutrophils [
103] with a key role for Sqstm1/p62 in autophagic control of infection [
104].