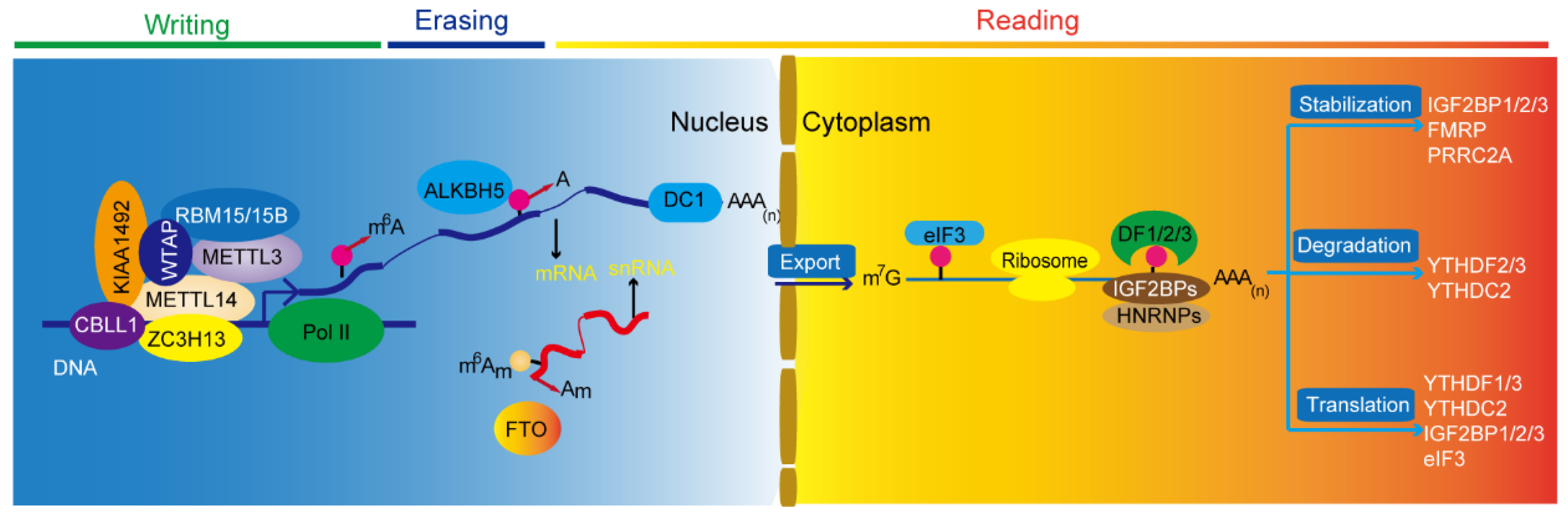
Glioblastoma is the most common and most lethal primary malignant brain tumor. N6-methyladenosine (m6A) is one of widespread and abundant internal messenger RNA (mRNA) modification found in eukaryotes. Accumulated evidence demonstrates that m6A modification is aberrantly activated in human cancers and is critical for tumorigenesis and metastasis. m6A modification is also strongly involved in key signaling pathways and is associated with prognosis in glioblastoma. Here, the researchers briefly outline the functions of m6A and its regulatory proteins, including m6A writers, erasers, and readers on the fate of RNA. The researchers also summarize the latest breakthroughs in this field, describe the underlying molecular mechanisms that contribute to the tumorigenesis and progression, and highlight the inhibitors targeting the factors in m6A modification in glioblastoma. Further studies focusing on the specific pathways of m6A modification could help identify biomarkers and therapeutic targets that might prevent and treat glioblastoma.
- m6A modification
- glioblastoma
- epigenetics
- methyltransferase
- demethylase
- inhibitor
1. Introduction
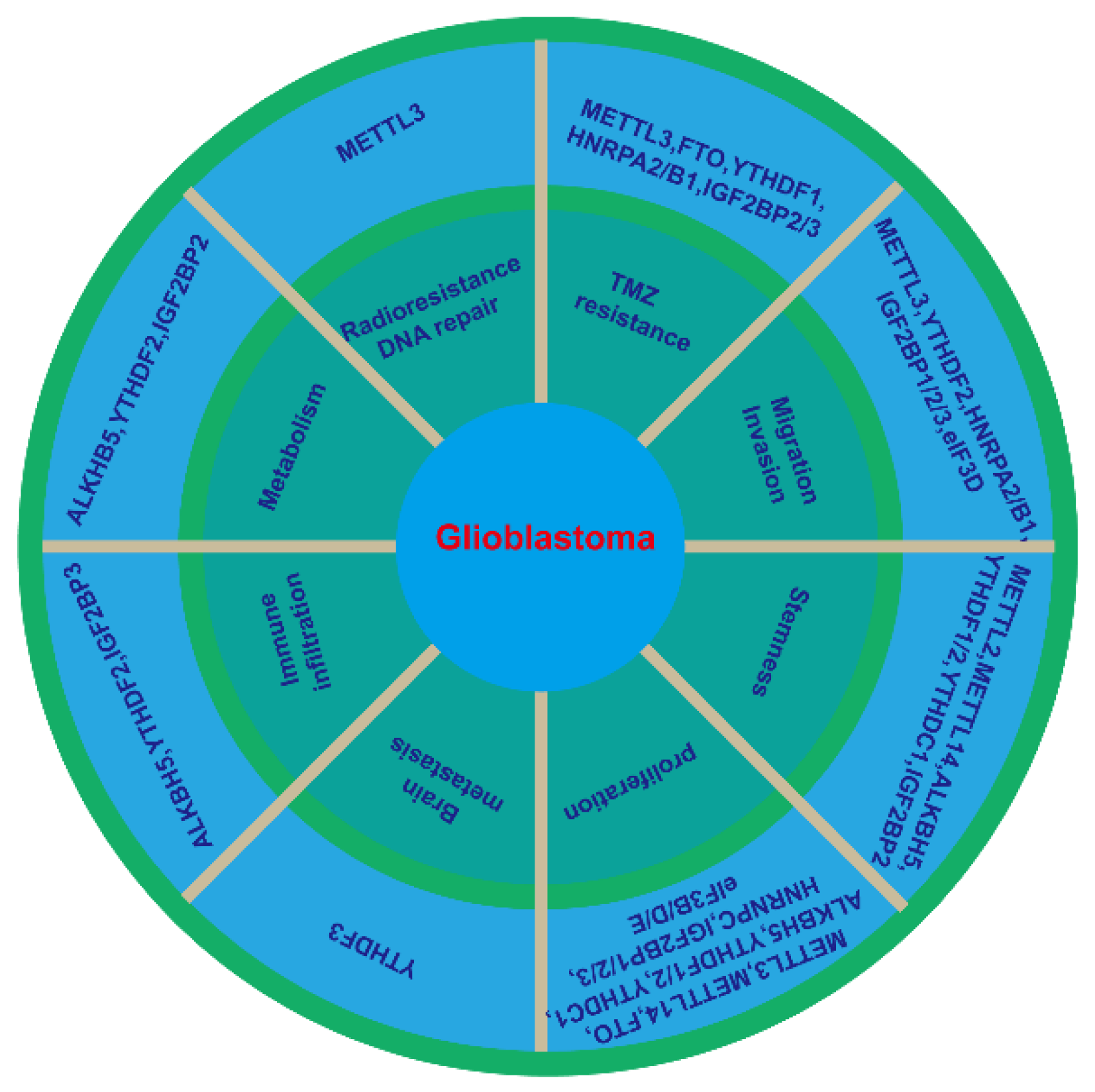
| Gene Name | Role in RNA Modification | Role in Glioblastoma | Mechanism | References |
|---|---|---|---|---|
| METTL3 | writer | oncogene | inhibiting sensitivity to γ-irradiation and enhancing DNA repair through recruitment of HuR to SOX2 mRNA. | [14] |
| oncogene | Activating NFκB in IDH-wildtype glioma after stabilization of MALAT1 | [15] | ||
| oncogene | Dysregulating the expression of epigenetically activated genes (the RNA editing, spicing and stability) | [16] | ||
| oncogene | Impairing the TMZ-sensitivity through m6A-modified DNA repair genes (MGMT and APNG)/EZH2 | [17][18] | ||
| Suppressor | Inhibiting epithelial to mesenchymal transition (EMT) and vasculogenic mimicry | [19] | ||
| Suppressor | promoting cell growth, cell differentiation, DNA damage response and cellular stress response by enhancing m6A | [20] | ||
| METTL14 | writer | Suppressor | promoting cell growth, cell differentiation, DNA damage response and cellular stress response by enhancing m6A | [20] |
| WTAP | writer | oncogene | A crucial interactor of the methyltransferase complex | [21][22] |
| FTO | eraser | oncogene | promoting proliferation and migration | [23] |
| oncogene | Increasing cell proliferation by targeting MYC transcripts | [24] | ||
| Suppressor | Inhibiting cell growth, migration and invasion by regulating m6A modification of primary pri-miR-10a processing | [25] | ||
| ALKBH5 | eraser | oncogene | Inhibiting cell proliferation and stemness through demethylating FOXM1 nascent transcripts and increasing HuR binding | [26] |
| oncogene | Demethylating G6PD transcript and enhancing its mRNA stability | [27] | ||
| oncogene | Enhancing hypoxia-induced TAM recruitment and immunosuppression by CXCL8/IL8 | [28] | ||
| oncogene | Increasing radioresistance by regulation homologous recombination | [29] | ||
| oncogene | Regulating TMZ resistance by promoting SOX2 expression | [30] | ||
| YTHDF1 | reader | oncogene | Promoting cell proliferation, stemness, and TMZ resistance via Musashi-1 | [31][32] |
| YTHDF2 | reader | oncogene | Positively correlating with immune cells markers, TAM markers and IDH1 | [33] |
| oncogene | Inhibiting cell proliferation, invasion and tumorigenesis through EGFR/SRC/ERK | [34] | ||
| oncogene | Accelerating UBXN1 mRNA degradation via METTL3-mediated m6A | [35] | ||
| oncogene | Linking epitranscriptomic modification by stabilizing MYC and VEGFA transcripts | [36] | ||
| YTHDF3 | oncogene | Promoting brain metastasis through enhancing the translation of m6A-mediated transcripts (ST6GALNAC5, GJA1 and EGFR) | [37] | |
| YTHDC1 | reader | oncogene | Promoting cell proliferation and stemness through VPS25-JAN-STAT | [38][39] |
| HNRNPC | reader | oncogene | Promoting cell proliferation, migration and invasion, and inhibiting apoptosis through Akt and p70S6K activation. | [40] |
| HNRPA2/B1 | reader | oncogene | Increasing cell viability, adhesion, migration, invasion, and TMZ resistance, and inhibiting apoptosis and ROS targeting STAT3, MMP-2/9 | [41][42] |
| IGF2BP1 | reader | oncogene | Targeted by non-coding RNAs and promoting cell proliferation, migration, and invasion | [43][44][45][46][47][48] |
| IGF2BP2 | reader | oncogene | Maintaining stemness and cell proliferation by regulating OXPHOS | [49] |
| oncogene | Targeted by non-coding RNAs and increasing TMZ resistance and proliferation | [50][51] | ||
| oncogene | Accelerating aerobic glycolysis by enhancing HK2 mRNA stability | [52] | ||
| oncogene | Promoting proliferation, and migration through recognition SRSF7 | [53] | ||
| oncogene | Promoting drug resitance by inhibition of PID1 through DANCR/FOCO1 axis | [54] | ||
| IGF2BP3 | reader | oncogene | Promoting proliferation, invasion and chemoresistance through PI3K and MAPK activation | [55] |
| oncogene | Targeted by miR-129-1 and miR-654 to induce proliferation and TMZ resistance | [56][57] | ||
| oncogene | Involving in macrophage infiltration in TME via stabilizing circNEIL3 | [58] | ||
| eIF3B | reader | oncogene | Promoting proliferation and inhibiting apoptosis | [59] |
| eIF3D | reader | oncogene | Promoting cell growth, colony formation and migration | [60] |
| eIF3E | reader | oncogene | Promoting proliferation through HIFs | [61] |
2. Potential Clinical Inhibitors of m6A Modification in Glioblastoma
3. Conclusions
This entry is adapted from the peer-reviewed paper 10.3390/biomedicines10050969
References
- Louis, D.N.; Perry, A.; Wesseling, P.; Brat, D.J.; Cree, I.A.; Figarella-Branger, D.; Hawkins, C.; Ng, H.K.; Pfister, S.M.; Reifenberger, G.; et al. The 2021 WHO Classification of Tumors of the Central Nervous System: A summary. Neuro-Oncology 2021, 23, 1231–1251.
- Huang, B.; Zhang, H.; Gu, L.; Ye, B.; Jian, Z.; Stary, C.; Xiong, X. Advances in Immunotherapy for Glioblastoma Multiforme. J. Immunol. Res. 2017, 2017, 3597613.
- Indraccolo, S.; Lombardi, G.; Fassan, M.; Pasqualini, L.; Giunco, S.; Marcato, R.; Gasparini, A.; Candiotto, C.; Nalio, S.; Fiduccia, P.; et al. Genetic, Epigenetic, and Immunologic Profiling of MMR-Deficient Relapsed Glioblastoma. Clin. Cancer Res. 2018, 25, 1828–1837.
- Desrosiers, R.; Friderici, K.; Rottman, F. Identification of methylated nucleosides in messenger RNA from Novikoff hepatoma cells. Proc. Natl. Acad. Sci. USA 1974, 71, 3971–3975.
- Schäfer, K.P. RNA Synthesis and Processing Reactions in a Subcellular System from Mouse L Cells. Hoppe-Seyler’s Z. Für Physiol. Chem. 1982, 363, 33–44.
- Meyer, K.D.; Saletore, Y.; Zumbo, P.; Elemento, O.; Mason, C.E.; Jaffrey, S.R. Comprehensive analysis of mRNA methylation reveals enrichment in 3′ UTRs and near stop codons. Cell 2012, 149, 1635–1646.
- Ke, S.; Alemu, E.A.; Mertens, C.; Gantman, E.C.; Fak, J.J.; Mele, A.; Haripal, B.; Zucker-Scharff, I.; Moore, M.J.; Park, C.Y.; et al. A majority of m6A residues are in the last exons, allowing the potential for 3′ UTR regulation. Genes Dev. 2015, 29, 2037–2053.
- Grozhik, A.V.; Linder, B.; Olarerin-George, A.O.; Jaffrey, S.R. Mapping m6A at Individual-Nucleotide Resolution Using Crosslinking and Immunoprecipitation (miCLIP). Methods Mol. Biol. 2017, 1562, 55–78.
- Dominissini, D.; Moshitch-Moshkovitz, S.; Schwartz, S.; Salmon-Divon, M.; Ungar, L.; Osenberg, S.; Cesarkas, K.; Jacob-Hirsch, J.; Amariglio, N.; Kupiec, M.; et al. Topology of the human and mouse m6A RNA methylomes revealed by m6A-seq. Nature 2012, 485, 201–206.
- Tuck, M.T. The formation of internal 6-methyladenine residues in eucaryotic messenger rna. Int. J. Biochem. 1992, 24, 379–386.
- Zaccara, S.; Ries, R.J.; Jaffrey, S.R. Reading, writing and erasing mRNA methylation. Nat. Rev. Mol. Cell Biol. 2019, 20, 608–624.
- He, L.; Li, H.; Wu, A.; Peng, Y.; Shu, G.; Yin, G. Functions of N6-methyladenosine and its role in cancer. Mol. Cancer 2019, 18, 176.
- Zhu, K.; Xie, V.; Huang, S. Epigenetic regulation of cancer stem cell and tumorigenesis. Adv. Cancer Res. 2020, 148, 1–26.
- Visvanathan, A.; Patil, V.; Abdulla, S.; Hoheisel, J.D.; Somasundaram, K. N6-Methyladenosine Landscape of Glioma Stem-Like Cells: METTL3 Is Essential for the Expression of Actively Transcribed Genes and Sustenance of the Oncogenic Signaling. Genes 2019, 10, 141.
- Chang, Y.Z.; Chai, R.C.; Pang, B.; Chang, X.; An, S.Y.; Zhang, K.N.; Jiang, T.; Wang, Y.Z. METTL3 enhances the stability of MALAT1 with the assistance of HuR via m6A modification and activates NF-κB to promote the malignant progression of IDH-wildtype glioma. Cancer Lett. 2021, 511, 36–46.
- Visvanathan, A.; Patil, V.; Arora, A.; Hegde, A.S.; Arivazhagan, A.; Santosh, V.; Somasundaram, K. Essential role of METTL3-mediated m6A modification in glioma stem-like cells maintenance and radioresistance. Oncogene 2017, 37, 522–533.
- Shi, J.; Chen, G.; Dong, X.; Li, H.; Li, S.; Cheng, S.; Li, Y.; Wang, L.; Yuan, J.; Qian, Z.; et al. METTL3 Promotes the Resistance of Glioma to Temozolomide via Increasing MGMT and ANPG in a m6A Dependent Manner. Front. Oncol. 2021, 11, 2849.
- Li, F.; Chen, S.; Yu, J.; Gao, Z.; Sun, Z.; Yi, Y.; Long, T.; Zhang, C.; Li, Y.; Pan, Y.; et al. Interplay of m6A and histone modifications contributes to temozolomide resistance in glioblastoma. Clin. Transl. Med. 2021, 11, e553.
- Tao, M.; Li, X.; He, L.; Rong, X.; Wang, H.; Pan, J.; Lu, Z.; Zhang, X.; Peng, Y. Decreased RNA m6A methylation enhances the process of the epithelial mesenchymal transition and vasculogenic mimicry in glioblastoma. Am. J. Cancer Res. 2022, 12, 893.
- Cui, Q.; Shi, H.; Ye, P.; Li, L.; Qu, Q.; Sun, G.; Sun, G.; Lu, Z.; Huang, Y.; Yang, C.-G.; et al. m6A RNA Methylation Regulates the Self-Renewal and Tumorigenesis of Glioblastoma Stem Cells. Cell Rep. 2017, 18, 2622–2634.
- Jin, D.-I.; Lee, S.W.; Han, M.-E.; Kim, H.-J.; Seo, S.-A.; Hur, G.-Y.; Jung, S.; Kim, B.-S.; Oh, S.-O. Expression and roles of Wilms’ tumor 1-associating protein in glioblastoma. Cancer Sci. 2012, 103, 2102–2109.
- Xi, Z.; Xue, Y.; Zheng, J.; Liu, X.; Ma, J.; Liu, Y. WTAP Expression Predicts Poor Prognosis in Malignant Glioma Patients. J. Mol. Neurosci. 2016, 60, 131–136.
- Li, F.; Zhang, C.; Zhang, G. m6A RNA Methylation Controls Proliferation of Human Glioma Cells by Influencing Cell Apoptosis. Cytogenet. Genome Res. 2019, 159, 119–125.
- Xiao, L.; Li, X.; Mu, Z.; Zhou, J.; Zhou, P.; Xie, C.; Jiang, S. FTO Inhibition Enhances the Antitumor Effect of Temozolomide by Targeting MYC-miR-155/23a Cluster-MXI1 Feedback Circuit in Glioma. Cancer Res. 2020, 80, 3945–3958.
- Zhang, S.; Zhao, S.; Qi, Y.; Li, B.; Wang, H.; Pan, Z.; Xue, H.; Jin, C.; Qiu, W.; Chen, Z.; et al. SPI1-induced downregulation of FTO promotes GBM progression by regulating pri-miR-10a processing in an m6A-dependent manner. Mol. Ther.-Nucleic Acids 2022, 27, 699–717.
- Zhang, S.; Zhao, B.S.; Zhou, A.; Lin, K.; Zheng, S.; Lu, Z.; Chen, Y.; Sulman, E.P.; Xie, K.; Bogler, O.; et al. m6A Demethylase ALKBH5 Maintains Tumorigenicity of Glioblastoma Stem-like Cells by Sustaining FOXM1 Expression and Cell Proliferation Program. Cancer Cell 2017, 31, 591–606.e6.
- Liu, Z.; Chen, Y.; Wang, L.; Ji, S. ALKBH5 Promotes the Proliferation of Glioma Cells via Enhancing the mRNA Stability of G6PD. Neurochem. Res. 2021, 46, 3003–3011.
- Dong, F.; Qin, X.; Wang, B.; Li, Q.; Hu, J.; Cheng, X.; Guo, D.; Cheng, F.-L.; Fang, C.; Tan, Y.; et al. ALKBH5 Facilitates Hypoxia-Induced Paraspeckle Assembly and IL8 Secretion to Generate an Immunosuppressive Tumor Microenvironment. Cancer Res. 2021, 81, 5876–5888.
- Kowalski-Chauvel, A.; Lacore, M.G.; Arnauduc, F.; Delmas, C.; Toulas, C.; Cohen-Jonathan-Moyal, E.; Seva, C. The m6A RNA Demethylase ALKBH5 Promotes Radioresistance and Invasion Capability of Glioma Stem Cells. Cancers 2020, 13, 40.
- Liu, B.; Zhou, J.; Wang, C.; Chi, Y.; Wei, Q.; Fu, Z.; Lian, C.; Huang, Q.; Liao, C.; Yang, Z.; et al. LncRNA SOX2OT promotes temozolomide resistance by elevating SOX2 expression via ALKBH5-mediated epigenetic regulation in glioblastoma. Cell Death Dis. 2020, 11, 384.
- Xu, C.; Yuan, B.; He, T.; Ding, B.; Li, S. Prognostic values of YTHDF1 regulated negatively by mir-3436 in Glioma. J. Cell. Mol. Med. 2020, 24, 7538–7549.
- Yarmishyn, A.A.; Yang, Y.-P.; Lu, K.-H.; Chen, Y.-C.; Chien, Y.; Chou, S.-J.; Tsai, P.-H.; Ma, H.-I.; Chien, C.-S.; Chen, M.-T.; et al. Musashi-1 promotes cancer stem cell properties of glioblastoma cells via upregulation of YTHDF1. Cancer Cell Int. 2020, 20, 597.
- Lin, X.; Wang, Z.; Yang, G.; Wen, G.; Zhang, H. YTHDF2 correlates with tumor immune infiltrates in lower-grade glioma. Aging 2020, 12, 18476–18500.
- Fang, R.; Chen, X.; Zhang, S.; Shi, H.; Ye, Y.; Shi, H.; Zou, Z.; Li, P.; Guo, Q.; Ma, L.; et al. EGFR/SRC/ERK-stabilized YTHDF2 promotes cholesterol dysregulation and invasive growth of glioblastoma. Nat. Commun. 2021, 12, 177.
- Chai, R.C.; Chang, Y.Z.; Chang, X.; Pang, B.; An, S.Y.; Zhang, K.N.; Chang, Y.H.; Jiang, T.; Wang, Y.Z. YTHDF2 facilitates UBXN1 mRNA decay by recognizing METTL3-mediated m6A modification to activate NF-κB and promote the malignant progression of glioma. J. Hematol. Oncol. 2021, 14, 109.
- Dixit, D.; Prager, B.C.; Gimple, R.C.; Poh, H.X.; Wang, Y.; Wu, Q.; Qiu, Z.; Kidwell, R.L.; Kim, L.J.Y.; Xie, Q.; et al. The RNA m6A Reader YTHDF2 Maintains Oncogene Expression and Is a Targetable Dependency in Glioblastoma Stem Cells. Cancer Discov. 2020, 11, 480–499.
- Chang, G.; Shi, L.; Ye, Y.; Shi, H.; Zeng, L.; Tiwary, S.; Huse, J.T.; Huo, L.; Ma, L.; Ma, Y.; et al. YTHDF3 Induces the Translation of m6A-Enriched Gene Transcripts to Promote Breast Cancer Brain Metastasis. Cancer Cell 2020, 38, 857–871.e7.
- Li, F.; Yi, Y.; Miao, Y.; Long, W.; Long, T.; Chen, S.; Cheng, W.; Zou, C.; Zheng, Y.; Wu, X.; et al. N6-Methyladenosine Modulates Nonsense-Mediated mRNA Decay in Human Glioblastoma. Cancer Res. 2019, 79, 5785–5798.
- Zhu, X.; Yang, H.; Zhang, M.; Wu, X.; Jiang, L.; Liu, X.; Lv, K. YTHDC1-mediated VPS25 regulates cell cycle by targeting JAK-STAT signaling in human glioma cells. Cancer Cell Int. 2021, 21, 645.
- Park, Y.M.; Hwang, S.J.; Masuda, K.; Choi, K.M.; Jeong, M.R.; Nam, D.H.; Gorospe, M.; Kim, H.H. Heterogeneous Nuclear Ribonucleoprotein C1/C2 Controls the Metastatic Potential of Glioblastoma by Regulating PDCD4. Mol. Cell. Biol. 2012, 32, 4237–4244.
- Deng, J.; Chen, S.; Wang, F.; Zhao, H.; Xie, Z.; Xu, Z.; Zhang, Q.; Liang, P.; Zhai, X.; Cheng, Y. Effects of hnRNP A2/B1 Knockdown on Inhibition of Glioblastoma Cell Invasion, Growth and Survival. Mol. Neurobiol. 2016, 53, 1132–1144.
- Li, L.; Wu, M.; Wang, C.; Yu, Z.; Wang, H.; Qi, H.; Xu, X. β-Asarone Inhibits Invasion and EMT in Human Glioma U251 Cells by Suppressing Splicing Factor HnRNP A2/B1. Molecules 2018, 23, 671.
- Li, Z.-W.; Xue, M.; Zhu, B.-X.; Yue, C.-L.; Chen, M.; Qin, H.-H. microRNA-4500 inhibits human glioma cell progression by targeting IGF2BP1. Biochem. Biophys. Res. Commun. 2019, 513, 800–806.
- Wang, R.-J.; Li, J.-W.; Bao, B.-H.; Wu, H.-C.; Du, Z.-H.; Su, J.-L.; Zhang, M.-H.; Liang, H.-Q. MicroRNA-873 (MiRNA-873) Inhibits Glioblastoma Tumorigenesis and Metastasis by Suppressing the Expression of IGF2BP1. J. Biol. Chem. 2015, 290, 8938–8948.
- Luo, Y.; Sun, R.; Zhang, J.; Sun, T.; Liu, X.; Yang, B. miR-506 inhibits the proliferation and invasion by targeting IGF2BP1 in glioblastoma. Am. J. Transl. Res. 2015, 7, 2007.
- Liu, P.; Zhao, P.; Li, B.; Xu, D.; Wang, K. LncRNA PCAT6 Regulated by YY1 Accelerates the Progression of Glioblastoma via miR-513/IGF2BP1. Neurochem. Res. 2020, 45, 2894–2902.
- Xue, J.; Zhong, S.; Sun, B.-M.; Sun, Q.-F.; Hu, L.-Y.; Pan, S.-J. Lnc-THOR silencing inhibits human glioma cell survival by activating MAGEA6-AMPK signaling. Cell Death Dis. 2019, 10, 866.
- Zhan, W.-L.; Gao, N.; Tu, G.-L.; Tang, H.; Gao, L.; Xia, Y. LncRNA LINC00689 Promotes the Tumorigenesis of Glioma via Mediation of miR-526b-3p/IGF2BP1 Axis. Neuromol. Med. 2021, 23, 383–394.
- Janiszewska, M.; Suvà, M.L.; Riggi, N.; Houtkooper, R.H.; Auwerx, J.; Clément-Schatlo, V.; Radovanovic, I.; Rheinbay, E.; Provero, P.; Stamenkovic, I. Imp2 controls oxidative phosphorylation and is crucial for preserving glioblastoma cancer stem cells. Genes Dev. 2012, 26, 1926–1944.
- Mineo, M.; Ricklefs, F.; Rooj, A.K.; Lyons, S.M.; Ivanov, P.; Ansari, K.I.; Nakano, I.; Chiocca, E.A.; Godlewski, J.; Bronisz, A. The Long Non-coding RNA HIF1A-AS2 Facilitates the Maintenance of Mesenchymal Glioblastoma Stem-like Cells in Hypoxic Niches. Cell Rep. 2016, 15, 2500–2509.
- Wang, X.; Li, X.; Zhou, Y.; Huang, X.; Jiang, X. Long non-coding RNA OIP5-AS1 inhibition upregulates microRNA-129-5p to repress resistance to temozolomide in glioblastoma cells via downregulating IGF2BP2. Cell Biol. Toxicol. 2021, 1–15.
- Liu, H.; Qin, S.; Liu, C.; Jiang, L.; Li, C.; Yang, J.; Zhang, S.; Yan, Z.; Liu, X.; Yang, J.; et al. m6A reader IGF2BP2-stabilized CASC9 accelerates glioblastoma aerobic glycolysis by enhancing HK2 mRNA stability. Cell Death Discov. 2021, 7, 292.
- Cun, Y.; An, S.; Zheng, H.; Lan, J.; Chen, W.; Luo, W.; Yao, C.; Li, X.; Huang, X.; Sun, X.; et al. Specific Regulation of m6A by SRSF7 Promotes the Progression of Glioblastoma. Genom. Proteom. Bioinform. 2021.
- Han, J.; Yu, X.; Wang, S.; Wang, Y.; Liu, Q.; Xu, H.; Wang, X. IGF2BP2 Induces U251 Glioblastoma Cell Chemoresistance by Inhibiting FOXO1-Mediated PID1 Expression Through Stabilizing lncRNA DANCR. Front. Cell Dev. Biol. 2022, 9, 659228.
- Suvasini, R.; Shruti, B.; Thota, B.; Shinde, S.V.; Friedmann-Morvinski, D.; Nawaz, Z.; Prasanna, K.V.; Thennarasu, K.; Hegde, A.S.; Arivazhagan, A.; et al. Insulin Growth Factor-2 Binding Protein 3 (IGF2BP3) Is a Glioblastoma-specific Marker That Activates Phosphatidylinositol 3-Kinase/Mitogen-activated Protein Kinase (PI3K/MAPK) Pathways by Modulating IGF-2. J. Biol. Chem. 2011, 286, 25882–25890.
- Kouhkan, F.; Mobarra, N.; Soufi-Zomorrod, M.; Keramati, F.; Rad, S.M.A.H.; Fathi-Roudsari, M.; Tavakoli, R.; Hajarizadeh, A.; Ziaei, S.; Lahmi, R.; et al. MicroRNA-129-1 acts as tumour suppressor and induces cell cycle arrest of GBM cancer cells through targeting IGF2BP3 and MAPK1. J. Med Genet. 2015, 53, 24–33.
- Jin, P.; Huang, Y.; Zhu, P.; Zou, Y.; Shao, T.; Wang, O. CircRNA circHIPK3 serves as a prognostic marker to promote glioma progression by regulating miR-654/IGF2BP3 signaling. Biochem. Biophys. Res. Commun. 2018, 503, 1570–1574.
- Pan, Z.; Zhao, R.; Li, B.; Qi, Y.; Qiu, W.; Guo, Q.; Zhang, S.; Zhao, S.; Xu, H.; Li, M.; et al. EWSR1-induced circNEIL3 promotes glioma progression and exosome-mediated macrophage immunosuppressive polarization via stabilizing IGF2BP3. Mol. Cancer 2022, 21, 16.
- Liang, H.; Ding, X.; Zhou, C.; Zhang, Y.; Xu, M.; Zhang, C.; Xu, L. Knockdown of eukaryotic translation initiation factors 3B (EIF3B) inhibits proliferation and promotes apoptosis in glioblastoma cells. Neurol. Sci. 2012, 33, 1057–1062.
- Ren, M.; Zhou, C.; Liang, H.; Wang, X.; Xu, L. RNAi-Mediated Silencing of EIF3D Alleviates Proliferation and Migration of Glioma U251 and U87MG Cells. Chem. Biol. Drug Des. 2015, 86, 715–722.
- Chai, R.-C.; Wang, N.; Chang, Y.-Z.; Zhang, K.-N.; Li, J.-J.; Niu, J.-J.; Wu, F.; Liu, Y.-Q.; Wang, Y.-Z. Systematically profiling the expression of eIF3 subunits in glioma reveals the expression of eIF3i has prognostic value in IDH-mutant lower grade glioma. Cancer Cell Int. 2019, 19, 155.
- Moroz-Omori, E.V.; Huang, D.; Bedi, R.K.; Cheriyamkunnel, S.J.; Bochenkova, E.; Dolbois, A.; Rzeczkowski, M.D.; Li, Y.; Wiedmer, L.; Caflisch, A. METTL3 Inhibitors for Epitranscriptomic Modulation of Cellular Processes. ChemMedChem 2021, 16, 3035–3043.
- Huff, S.; Tiwari, S.K.; Gonzalez, G.M.; Wang, Y.; Rana, T.M. m6A-RNA Demethylase FTO Inhibitors Impair Self-Renewal in Glioblastoma Stem Cells. ACS Chem. Biol. 2021, 16, 324–333.
- Malacrida, A.; Rivara, M.; Di Domizio, A.; Cislaghi, G.; Miloso, M.; Zuliani, V.; Nicolini, G. 3D proteome-wide scale screening and activity evaluation of a new ALKBH5 inhibitor in U87 glioblastoma cell line. Bioorg. Med. Chem. 2020, 28, 115300.
- Selberg, S.; Seli, N.; Kankuri, E.; Karelson, M. Rational Design of Novel Anticancer Small-Molecule RNA m6A Demethylase ALKBH5 Inhibitors. ACS Omega 2021, 6, 13310–13320.
- Wallis, N.; Oberman, F.; Shurrush, K.; Germain, N.; Greenwald, G.; Gershon, T.; Pearl, T.; Abis, G.; Singh, V.; Singh, A.; et al. Small molecule inhibitor of Igf2bp1 represses Kras and a pro-oncogenic phenotype in cancer cells. RNA Biol. 2021, 19, 26–43.
- Li, Y.; Li, Y.; Ge, P.; Ma, C. MiR-126 Regulates the ERK Pathway via Targeting KRAS to Inhibit the Glioma Cell Proliferation and Invasion. Mol. Neurobiol. 2016, 54, 137–145.