Hybrid rye breeding leads to considerably higher grain yield and a higher revenue to the farmer. The basis of hybrid seed production is the CMS-inducing Pampa (P) cytoplasm derived from an Argentinean landrace and restorer-to-fertility (Rf) genes. Breeding is based on inbred line development and intensive testing for line per se performance and general combining ability (GCA). The finally selected inbred lines are used to compose a topcross hybrid with two genetically different seed parent lines (A-P, B-N) and a restorer synthetic composed of two S2 lines (SynRf). European restorer sources show low-to-moderate pollen-fertility levels. This results in higher susceptibility to ergot (Claviceps purpurea) because rye pollen and ergot spores are in strong competition for the unfertilized stigma. Rf genes from non-adapted Iranian primitive rye and old Argentinean cultivars proved to be most effective. The major Rf gene in these sources was localized on chromosome 4RL, which is also a hotspot of restoration in other Triticeae. Commercial hybrids with these Rf genes showed a similar low ergot infection when compared with population cultivars. The great future challenges of climate change, such as increased drought stress tolerance, improved lodging tolerance, as well as the increasing need for resistant varieties can effectively only be met by hybrid breeding.
- Claviceps purpurea
- CMS
- hybrid breeding
- pollen
- restorer genes
- yield
1. Basics of hybrid rye breeding
Hybrid rye breeding leads to considerably higher grain yield and a higher revenue to the farmer. The basis of hybrid seed production is the cytoplasmic-male sterility (CMS)-inducing Pampa (P) cytoplasm derived from an Argentinean landrace[5] and restorer-to-fertility (Rf) genes[6]. To ensure a maximal heterosis, two gene pools are used, that are in Central Europe the Petkus (seed parent) and the Carsten (pollen parent) gene pool. The pools were derived from two breeding companies that used different landraces for starting their selection work in the 19th and the beginning 20th century. The basis of hybrid breeding are inbred lines that are produced in rye by subsequent selfing with the use of a dominant single-gene mutation leading to full self-fertility (sf).
Breeding starts with crossing of parents (Figure 1), followed by two selfings of the progenies and subsequent per se performance test of S2 lines for highly heritable traits (e.g., vitality, heading date, plant height, disease resistances, lodging tolerance, 1000-grain weight, falling number). The selected fraction is crossed in the seed parent pool to a CMS donor to start the backcrossing procedure that is necessary to result at the end in near-isogenic lines with the CMS-inducing P cytoplasm and the normal cytoplasm. During backcrossing selection for non-restorer is performed. In contrast, in the pollen parent pool a strict selection for Rf genes has to practised. Here, only two selfings are done, because afterwards the best selected lines are crossed to a synthetic. In parallel to selfing and backcrossing two stages of yield tests (YT) are made by crossing the inbred lines with testers from the opposite pool. These testcrosses (TC) are planted in 3-5 locations in two replicates to select for general combining ability (GCA) for grain yield and the other mentioned traits. The finally selected inbred lines are used to compose a topcross hybrid with two genetically different seed parent lines (A-P, B-N) and a restorer synthetic composed of two S2 lines (SynRf). This complex hybrid is necessary due to the high inbreeding depression that rye is suffering as an obligate cross-pollinating crop.
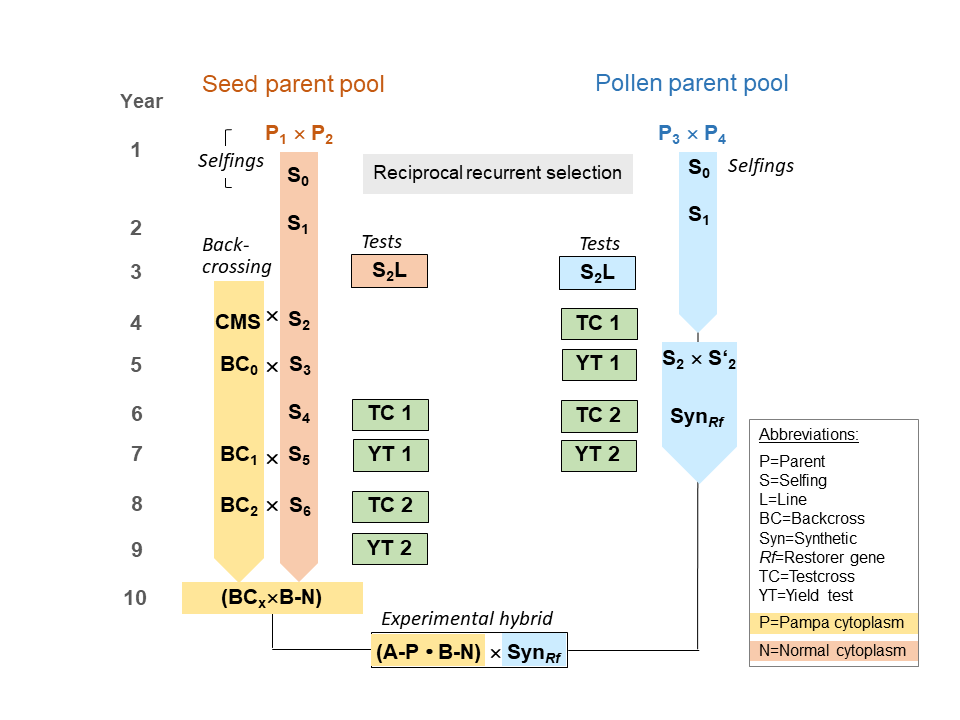
Figure 1. Scheme for hybrid rye breeding
Hybrid rye production requires strict logistics for seed multiplication, because rye is an effective wind pollinator. All stands of seed production must be strictly isolated and checked for outcrossings several times. Mostly the line multiplications (A-P × A-N, B-N) and the production of basic seed (A-P × B-N) are done in foreign countries with no commercial rye growing (France, Italy, Hungary). The lines are planted in stripes with a 3:1 proportion for females:males. The multiplication of the synthetics and the production of certified seed (A-P • B-N × SynRf) is done in the area of marketing of the rye hybrid cultivars. The certified seed is produced in a technical mixture of 90-95% CMS seed parent and 5-10% restorer synthetic. This approach cheapens production, but currently requires the presence of synthetics to escape the strong inbreeding depression of lines.
2. Ergot as a challenge for hybrid rye production
Since Medieval times, the results of ergot infections have been documented. Rye and ergot were so closely interwoven that botanical drawings of rye in the 17th century included ergot. Ergot refers to the blackish purple sclerotia of the fungus Claviceps purpurea (Fries ex Fries) Tulasne that extend far beyond the husks of rye (Figure 2a). They contain more than 80 ergot alkaloids, of which six alkaloids and their corresponding epimers were found to be the most important ones in Europe. Prolonged consumption of ergot-containing flour, which was common among the peasant population in the Middle Ages, leads to a serious disease, ergotism, that is characterized either by nervous disorders (fever, muscle spasms, tremor, paralysis, hallucinations) or by vasoconstriction effects (gangrene, violent burning, shooting pain in the fingers and toes, called “St. Anthony’s fire”).

All mammalian livestock are affected by ergot contamination of feed. Epidemic outbreaks in the Middle Ages resulted in a death rate of between 10% and 20% and a reduction in the birth rate due to the premature onset of labor and miscarriages[7]. The main factor for ergot epidemics is rainy weather during rye flowering. High humidity stimulates fungal growth and reduces the availability of rye pollen. The fungus cannot grow through intact glumes and the spores must be brought into the open florets by insects or by aerosols induced by rain and wind. Therefore, cross-pollinating grasses, such as rye, are a primary target of the fungus. The infection success of the fungus is reduced when the florets are already pollinated and the then-closed glumes prevent access to the stigma. Ergot spores mimic the fertilization process of rye. After landing on the stigma, the germ tubes grow through the pistil, like the pollen tubes, and infect the ovary exclusively. Therefore, the pollination has a considerable effect on infection frequency and disease development[8]. The combination of less available pollen, poor pollen flight properties, and an extended opening time of the unfertilized florets in rainy weather gives the fungus a competitive advantage in relation to pollen. This is less pronounced as the cultivar sheds more pollen (Figure 2b).The close association between pollen availability and ergot susceptibility poses a problem for hybrid breeding when full pollen shedding cannot be guaranteed due to lack of superior restoration. In the early hybrid cultivars, pollen-fertility restoration was inferior, leading to high ergot contamination when the weather was conducive for the disease. From 1993 to 2004, the mean percentage of natural ergot incidence in the German rye harvest was 0.04% for population cultivars and 0.18% for hybrid cultivars according to results from the Max-Rubner Institute, Detmold, Germany. This was not acceptable, as the EU tolerance limit for ergot sclerotia in commercial grain lots for food at that time was 0.05%, and for feed it was 0.1%. The high susceptibility of the early hybrids to ergot stimulated research for better restorer (Rf) genes.
European restorer sources show an oligogenic inheritance, with major and minor Rf genes, and mostly result in low-to-moderate pollen-fertility levels. This results in higher susceptibility to ergot because rye pollen and ergot spores are in strong competition for the unfertilized stigma. Rf genes from non-adapted Iranian primitive rye and old Argentinean cultivars proved to be most effective. The major Rf genes in these sources were all localized on chromosome 4RL, which is also a hotspot of restoration in other Triticeae. Marker-based introgression into elite rye materials led to a yield penalty and taller progenies. The Rfp1 gene from the primitive rye IRAN IX was fine-mapped, and two linked genes of equal effects were detected. Commercial hybrids with this gene showed a similar low ergot infection when compared with population cultivars.
3. How to Proceed in Future Breeding
Hybrid breeding is the future of rye! The higher yield performance allows the crop to compete with wheat, which usually fetches a higher price on the market. In view of climate change, with drier and warmer conditions in the future, hybrid rye is superior in all growing regions. It also requires less pesticide and nitrogen fertilization, in line with the European Green Deal, which calls for higher environmental protection standards in agriculture. The EU has significantly tightened the limits for ergot since 1 January 2022. In addition to the previous limit of a maximum of 0.5 g/kg sclerotia and sclerotial fragments on unprocessed rye, harvested grain may not contain more than 500 µg/kg total alkaloids on rye milling products for human consumption. From 1 July 2024, the limits will be reduced to 0.2 g/kg sclerotia and a maximum of 250 µg/kg alkaloids according to the EU Commission Regulation 2021/1399. Therefore, the high pollen-shedding of hybrid varieties must be the top priority to avoid ergot.
Besides maximizing pollen-shedding by using non-adapted Rfp genes, the question of resistance of rye to the pathogen Claviceps purpurea, arises. It was previously taken for granted that ergot infection does not cause any defense reactions in rye. However, recent transcriptomic studies showed that in both wheat and rye, the host plant responds to the infection by upregulating defense-related genes, precisely when the germ tube reaches the ovary[9]. Thus, the host obviously recognizes the Claviceps infection. Indeed, it has been shown several times that diverse sets of both fully pollen-fertile rye and fully male-sterile rye, when grown under pollen-isolated conditions, showed small but significant differences in their ergot reaction[10]. These quantitative maternal differences could be exploited by targeted selection, as shown by some commercial hybrids without Rfp1 gene and with a low restorer index, but also lower ergot severity after artificial infection (Figure 3). The best-restored hybrids with Rfp1 have a similar low ergot severity like population cultivars. In Germany, Poland, and Austria, the official cultivar registration procedure includes multi-locational testing for ergot severity over two years, with artificial inoculation in a special experimental setting.
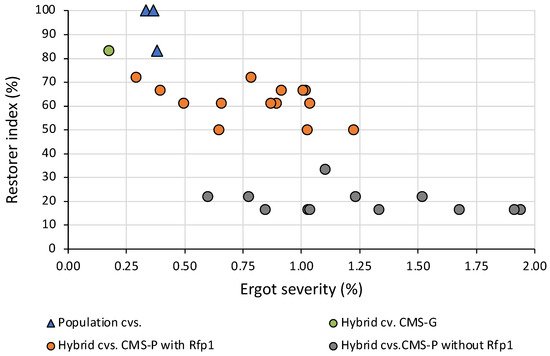
The high effectiveness of non-adapted Rfp genes will play a prominent role in future hybrid breeding. The main task is to reduce the obvious yield penalty and other negative agronomic features. Even if the introgressed genomic section is significantly shortened, a significant yield loss remains that could not be further reduced using more closely linked markers. However, the effects of the same Rfp gene on different genotypes significantly differs. Therefore, a consequent selection in the elite gene pools for mitigating undesired side effects of Rfp genes seems feasible in the long run. This would replicate the coevolution that has occurred in adapted CMS/Rf systems over evolutionary time periods.
This coevolution could be enhanced in a breeding context, when environmentally stable adaptation genes exist in the elite gene pools that lower the negative agronomic effects of non-adapted Rf genes. Research activities should be started to identify and to exploit these genes. In conclusion, the development of commercial hybrid cultivars in P cytoplasm, with a similarly low or an even lower ergot severity than population cultivars, appears to be promising.
In hybrid rye breeding, modern genomic techniques are practised to increase the performance of hybrids and ensure a steady selection gain in future (Table 1). Marker-assisted selection is useful for all monogenically inherited traits. Marker-assisted backcrossing helps to shorten the backcrossing procedure in the seed-parent pool (rf. to Figure 1) by selecting against the CMS donor background. Using a low-density marker array in BC3 already the same low donor background could be achieved like normally in BC6. QTL mapping is an important tool when lines with special alleles are searched that are not available in the current breeding populations. With this tool, the genetic architecture of the trait can be analysed with high power, because even rare alleles are pushed to an allele frequency of 50% in bi-parental crosses.
Table 1. Genome-based techniques and their use in hybrid rye breeding (KASP=kompetitive allele-specific PCR)
|
Technique |
Useful for … |
Requirement |
|
Marker-assisted selection (MAS) |
Introgression of mono-genic resistances, Rf genes, dwarfing genes |
Closely linked diagnostic KASP markers |
|
Marker-assisted backcrossing (MAB) |
Conversion of N lines: |
Low-density marker array |
|
Quantitative trait loci (QTL) mapping |
Detection of rare alleles with large effects |
Low-density marker array |
|
Genome wide association mapping (GWAS) |
Detection of common alleles with smaller effects |
Medium-density marker array |
|
Genomic selection (GS) |
Selection of genome-wide marker effects for all traits |
High-density marker array |
Genomic selection (GS) is already practised in some breeding companies. Typically, GS is done after the first S2 line per se performance (rf. to Fig. 1). Based on previous cycles that are used as training population, superior lines in GCA for grain yield and other traits are predicted solely on their genomic composition and only a reduced set of positively predicted lines enters the costly 2-year testcross procedure[11]. After the yield test, a final selection can take place based on an index of the phenotypically measured and genomically predicted GCA values. Implementing GS allows to test about 50% more candidates with the same budget, thus considerably increasing selection gain.
The great future challenges of climate change, such as increased drought stress tolerance, improved lodging tolerance, as well as the increasing need for resistant varieties can only be met effectively and as quickly as possible with hybrid breeding.
This entry is adapted from the peer-reviewed paper 10.3390/plants11091115
References
- Karl-Ernst Behre; The history of rye cultivation in Europe. Vegetation History and Archaeobotany 1992, 1, 141-156, 10.1007/bf00191554.
- FAOSTAT . FAO. Retrieved 2022-5-5
- Geiger, H.H.; Miedaner, T. Rye Breeding; Carena, M.J., Eds.; Springer: New York, USA, 2009; pp. 157-181.
- Friedrich Laidig; Hans-Peter Piepho; Dirk Rentel; Thomas Drobek; Uwe Meyer; Alexandra Huesken; Breeding progress, variation, and correlation of grain and quality traits in winter rye hybrid and population varieties and national on-farm progress in Germany over 26 years. Theoretical and Applied Genetics 2017, 130, 981-998, 10.1007/s00122-017-2865-9.
- H. H. Geiger; F. W. Schnell; Cytoplasmic Male Sterility in Rye ( Secale cereale L.) 1. Crop Science 1970, 10, 590-593, 10.2135/cropsci1970.0011183x001000050043x.
- Geiger, H.H.; Restoration of pollen fertility to cytoplasmic male sterile rye. Theor. Appl. Genet. 1972, 42, 32-33, .
- Pieter W.J. van Dongen; Akosua N.J.A. de Groot; History of ergot alkaloids from ergotism to ergometrine. European Journal of Obstetrics & Gynecology and Reproductive Biology 1995, 60, 109-116, 10.1016/0028-2243(95)02104-z.
- Thomas Miedaner; Hartwig H. Geiger; Biology, Genetics, and Management of Ergot (Claviceps spp.) in Rye, Sorghum, and Pearl Millet. Toxins 2015, 7, 659-678, 10.3390/toxins7030659.
- Birgitt Oeser; Sabine Kind; Selma Schurack; Thomas Schmutzer; Paul Tudzynski; Janine Hinsch; Cross-talk of the biotrophic pathogen Claviceps purpurea and its host Secale cereale.. BMC Genomics 2017, 18, 273, 10.1186/s12864-017-3619-4.
- Anna Kodisch; Brigitta Schmiedchen; Jakob Eifler; Andres Gordillo; Dörthe Siekmann; Franz Joachim Fromme; Michael Oberforster; Thomas Miedaner; Maternal differences for the reaction to ergot in unfertilized hybrid rye (Secale cereale). European Journal of Plant Pathology 2022, 163, 181-191, 10.1007/s10658-022-02467-0.
- Wilde, P.; Miedaner, T. Hybrid rye breeding ; Rabanus-Wallace, M.T., Stein, N., Eds.; Springer: Cham, 2021; pp. 13-41.