The primary cilium is a small microtubule-based organelle that extends from the apical surface of most eukaryotic cells into the extracellular space for sensing and transducing a wide range of cues. Defects in cilia growth and function are associated with a group of human inherited multisystemic diseases, known as ciliopathies. In recent years a rising number of ciliary proteins have been described at extraciliary sites, both in ciliated and non-ciliated cells, and have been implicated in cilium-independent functions and different cellular processes. Hematopoietic cells, including T lymphocytes, do not form primary cilia. However, non-ciliated T cells co-opt the ciliogenesis machinery for the assembly and function of the immunological synapse, a well-organized structure formed by immune cells – multiple types of T cells, mast cells, NK cells, B cells, neutrophils, macrophages, and dendritic cells – allowing for antigen recognition and signaling, information exchange and polarized release of molecules into the synaptic cleft. The identification of many, unexpected similarities between the primary cilium and the T cell immunological synapse at the structural, functional and molecular levels have highlighted the homology between these structures, even though they show disparate morphologies.
- primary cilium
- proteins
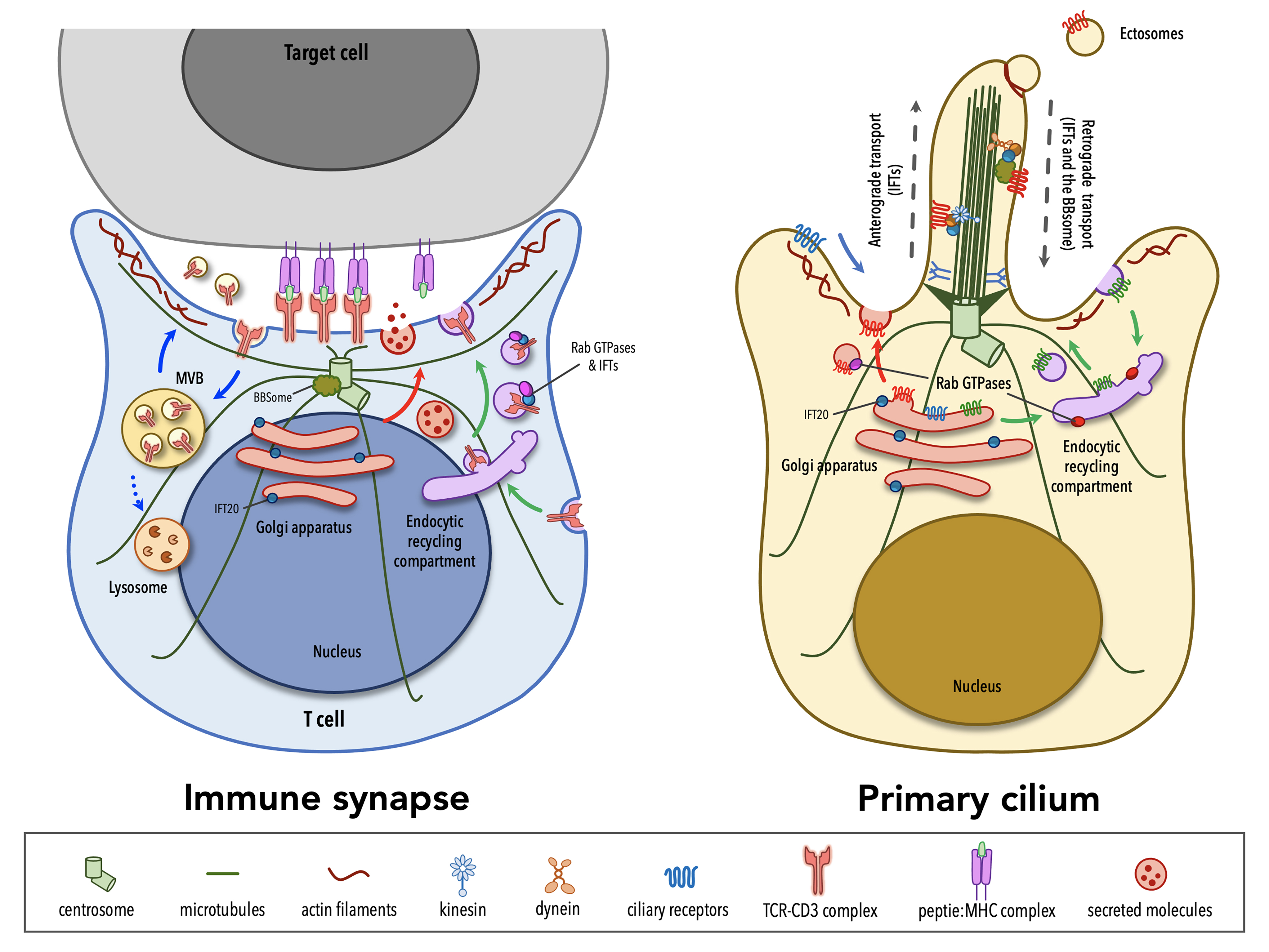
1. An overview of the immunological synapse and the primary cilium
The immunological synapse (IS) is a specialized, adhesive junction that forms between either immune cells (e.g. a T helper cell and a B cell), or an immune cell and a different cell type (e.g. a cytotoxic T cell and its cognate target). This structure is characterized by: (i) close apposition of cell membranes that seals off a nanoscale space, called the synaptic cleft; (ii) cell-cell adhesion mediated by integrins; (iii) junction stability relative to the highly dynamic behavior of T lymphocytes in secondary lymphoid tissues; (iv) directed secretion of soluble factors (i.e. cytokines and cytotoxic molecules) in response to TCR signaling[1]. In the 1970s puzzling evidence highlighted physical interactions between immune cells as required for T cell help and cytotoxicity[2] and the relevance of this interaction to T cell activation and function was synthetized by Norcross as “synaptic basic for T cell activation”[3]. However, only in 1998 Kupfer took advantage of the rapid advances in fluorescence microscopy and molecular immunology to carry out a 3D analysis of T cell-APC conjugates, revealing the spatial organization of receptors and adhesion molecules in distinct SupraMolecular Activation Clusters within the IS. The mature bull’s eye synapse consists of three concentric regions[4]: a central SMAC (cSMAC) enriched in clustered T cell antigen receptors (TCRs) and the co-stimulatory receptor CD28, a peripheral SMAC (pSMAC) where adhesion molecules, such as LFA-1, are accumulated allowing for a tight adhesion between the cells and a lamellipodium-like distal SMAC (dSMAC).
The primary cilium is a single, non-motile projection that emerges from the surface of most cell types with some variations depending on cell cycle and differentiation stages[5]. First observed in the 19th century by Zimmermann[6], the primary cilium has long been considered an organelle that had lost its mobility function, and therefore a rudimental organelle, due to its 9+0 axoneme lacking the central microtubule pair and dynein arms. The fact that mutations in genes encoding for primary cilium components are cause of a wide range of human diseases, collectively known as ciliopathies[7], has led to rethink the primary cilium function. An in-depth study of ciliopathy-associated phenotypes has highlighted the signaling function of the primary cilium as well as the mechanisms and the regulators that control its assembly and maintenance.
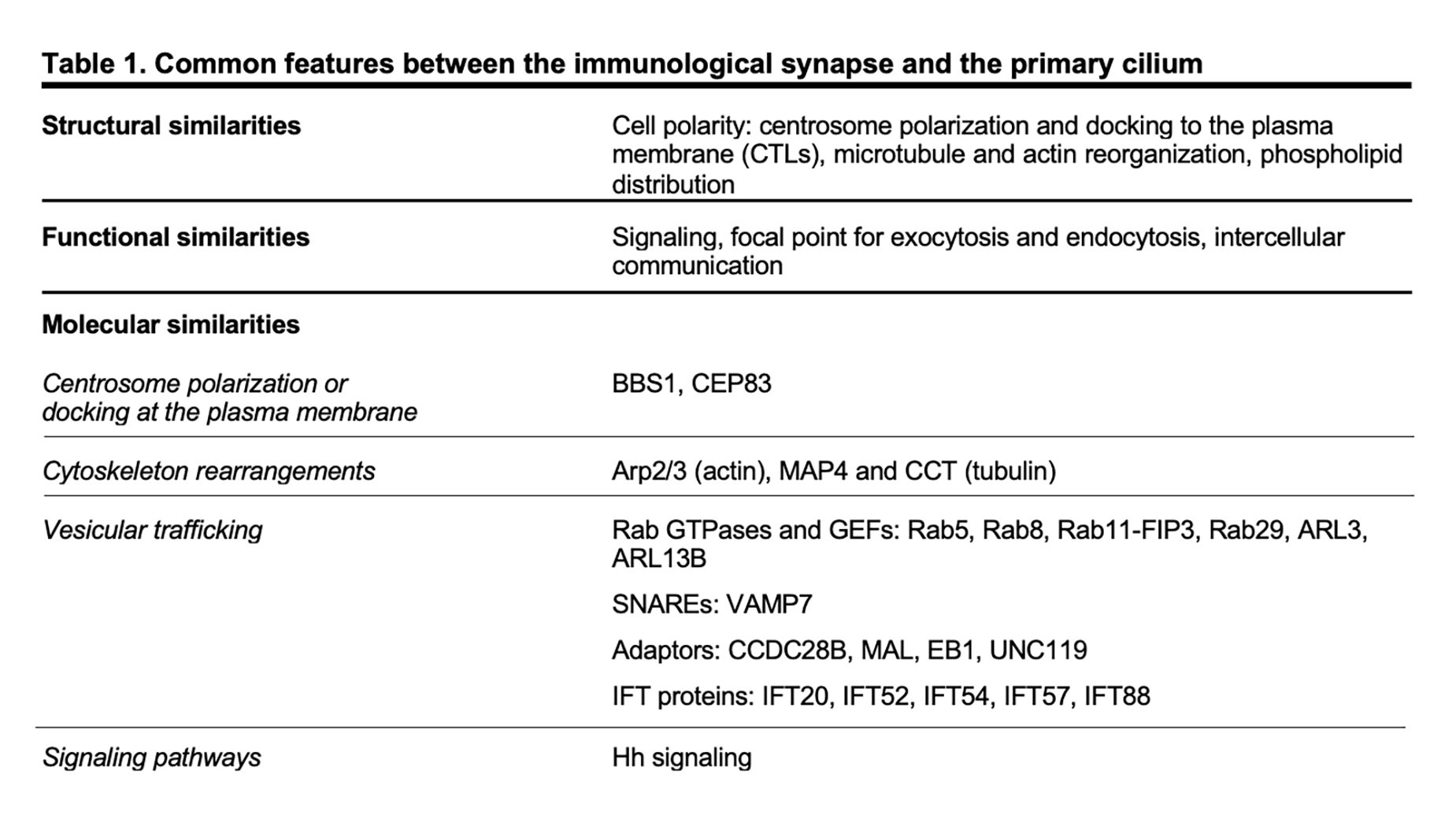
Although lymphocytes are one of the few cell types that lack primary cilia[8][9], electron micrographs of the IS showed that the centrosome docks at the synaptic membrane in CTLs, which is similar to the positioning of the basal body in the primary cilium, suggesting that these cells are able to form a “frustrated cilium” with centrosome docking at the synaptic membrane but not cilium elongation[10][11]. Additionally, T lymphocytes were found to express Intraflagellar Transport (IFT) proteins[12], which are essential for cilium formation and function, and to exploit them for vesicular trafficking and IS assembly. In subsequent years, several structural, functional and molecular parallels have revealed striking similarities between the IS and the primary cilium (Table 1), leading to the hypothesis that these structures might be functional homologues.
2. Structural similarities
The primary cilium and the IS are prime examples of polarized structures, the formation of which is concomitant with an asymmetric re-organization of cytoskeleton, cytoplasmic and plasma membrane-associated molecules, as well as intracellular organelles.
A hallmark of cell polarity is the reorientation of the centrosome, the most common microtubule-organizing center (MTOC)[13], that moves towards the apical side of the cell and eventually establishes a physical contact with the plasma membrane. The centrosome consists of two centrioles, the mother centriole endowed of distal and subdistal appendages and the daughter centriole lacking appendages, surrounded by a pericentrosomal material (PCM)[14]. An early event in ciliogenesis is the centrosome-to-basal-body conversion[15], a process that in some contexts relies on vesicular trafficking. Indeed, vesicles originating at the Golgi complex or at the recycling compartment accumulate on the top of the mother centriole to form a capping vesicle that moves in association with the mother centriole and fuses with the plasma membrane once it reaches the surface. The basal body docks via the distal appendages of the mother centriole to a portion of plasma membrane, which acquires some specialization features to become the ciliary membrane[16][17][18][19]. The polymerization of nine doublets of microtubules drives axoneme elongation, giving rise to a protrusion that emerges from the cell surface. In immune cells, the centrosome re-posits just beneath the synaptic membrane and only in the case of cytotoxic T cells (CTL) the mother centriole’s distal appendages have been observed to anchor the centrosome to the plasma membrane, allowing for the microtubule-mediated transport of lytic granules and their subsequent release to kill a target cell[10][11]. Centrosome polarization is mainly driven by cytosokeleton reorganization, even though a minor contribution of vesicular trafficking to this process cannot be ruled out as witnessed by the presence of a capping vesicle, which resembles that one of ciliated cells, over one of the centrioles in CD4+ T cells[20]. Of note, centrosome polarization does not only orient the microtubule array that originates from it, but also dictates the recruitment of other organelles, including the Golgi apparatus, and of vesicles that use microtubules as tracks for their directional, molecular-mediated movement towards the base of the primary cilium or in the IS proximity[21][22][10].
In addition to changes in the general microtubule organization, rearrangements of the actin cytoskeleton at the cell cortex occur during the early stages of ciliogenesis and IS assembly. A combination of proteomic and microscopy approaches has identified the presence of actin-binding proteins in the ciliary compartment[23][24][25][26][27][28], highlighting the importance of a more in-depth comprehension of actin cytoskeleton dynamics in the formation of primary cilia. Genetic depletion and inhibition studies, in which the turnover of actin filaments is tilted towards depolymerization, suggested that F-actin nucleation prevents cilia formation[29][30][31][32]. However, the role of the actin cytoskeleton in ciliogenesis is still controversial. In fact, more recent evidence has involved actin-binding proteins in ciliogenesis and cilia maintenance[33]. Remodeling of cortical actin is well described in T lymphocytes. The initial interaction between a T lymphocyte and its cognate cell results in the accumulation of actin at the contact site[34][35][36], followed by the reorganization of cortical actin in three discrete actin networks surrounding a central zone, which is relatively free of actin[37]: from the edge to the center, a lamellipodial branched actin network, a lamellar acto-myosin network, and actin foci that are thought to be associated with invadosome-like protrusions (ILPs). Beyond the dynamics of cortical actin, a common feature in ciliated and in non-ciliated cells is that the centrosome moves towards a hypodense actin region[38][11], where it has access to the plasma membrane for docking. Furthermore, an emerging role for a centrosomal pool of filamentous actin in ciliogenesis and IS assembly has been recently described. In ciliated cells, the transport of periciliary vesicles from the pericentrosomal region to the distal appendages of the mother centriole for primary cilium growth is mediated by the actin-based motor protein myosin-Va[39]. On the contrary, in non-ciliated T cells a local depletion of centrosomal F-actin is required for centrosome detachment from the nuclear envelope, allowing for its repositioning to the IS[40][41].
Consistent with their signaling function, both the IS and the primary cilium membrane show a specialized composition, enriched in receptors and signaling mediators, as well as in lipid rafts and phospholipids. Specifically, phospholipid distribution within the ciliary membrane is very similar to that one across the synapse, with an accumulation of phosphatidylinositol 4-phosphate PI(4)P in both ciliary and synaptic membrane that is replaced by phosphatidylinositol 4,5-biphosphate (PI(4,5)P2) and phosphatidylinositol 3,4,5-thipshosphate (PI(3,4,5)-P3) at the base of the cilium and at the edge of the IS[42][43][44]. A local accumulation of PI(4,5)P2, a lipid that is known to interact with actin-regulating proteins[45], at the tip of the primary cilium is a peculiarity of primary cilia, where both the release of ectosomes and the loss of the ciliary tips itself through an unusual form of cilia decapitation have been described as actin-mediated processes[46][47].
3. Functional similarities
Both the primary cilium and the IS have been extensively characterized as signaling devices. Cells engaged in a tight junction for minutes to hours exchange information via chemical signals (i.e. membrane-associated receptors and soluble molecules) and physical forces that are generated via the spatiotemporal organization and dynamics of the cell cytoskeleton. Each cell integrates information coming from different signaling pathways and produces appropriate biochemical and genetic responses. Although historically regarded as a vestigial organelle, it has become clear that the primary cilium can sense a variety of extracellular stimuli and transducing these incoming signals into a cellular response with relevant outcomes in multiple cellular and developmental processes in vertebrates[48][49]. For this reason, the primary cilium is now referred to as “cell antenna”.
In addition to and in many instances related to signaling, both the IS and the primary cilium are vesicular trafficking hotspots. This is the consequence of the fact that, as the primary cilium and the IS assemble, the Golgi apparatus and the recycling/endosomal compartment polarize just beneath the base of the cilium and the synaptic membrane, respectively. At the base of the primary cilium the periciliary membrane invaginates to form a cytoplasmatic invagination called the ciliary pocket (CiPo), acting as a specialized site for localized endocytosis and exocytosis of vesicles that accompany the delivery and retrieval of ciliary components to and from the primary cilium[50][51]. Several studies have demonstrated that cilia growth and function require the directional delivery of vesicles carrying lipids and some ciliary membrane proteins from the Golgi or recycling endosomes to the periciliary membrane region, where vesicles undergo exocytosis and lipids are incorporated into the ciliary membrane[52]. Alternatively, some ciliary membrane proteins can enter the ciliary membrane from the plasma membrane through lateral diffusion[53][54][55][56]. Protein trafficking within the ciliary compartment is mediated by IFT particles, each comprising the IFT-A and IFT-B subcomplexes, which move along axonemal microtubules in association with the molecular motors kinesin-2 and cytoplasmic dynein-2[57][58]. The termination of many signaling pathways triggered at the primary cilium is regulated through the clearance of specific receptors from the ciliary compartment[59][60] and some of these are internalized at the ciliary pocket through clathrin-dependent endocytosis[48]. When the transport of activated ciliary receptors from cilia back into the cell fails, they can be incorporated into ectosomes that bud from the tip of the primary cilium with the assistance of newly polymerized actin filaments[46]. Some studies have demonstrated that the primary cilium is able not only to receive incoming signals, but also to produce signals that are transferred to other cells via extracellular vesicles (EVs)[61]. The functions of EVs released at the primary cilium are still poorly investigated. Recent evidence suggests the release of EVs as a means for a rapid modulation of the protein composition of the ciliary compartment, with relevant implications in signaling[46][62]. Although the role of ciliary EVs in intercellular communication is only starting to be experimentally tested, ciliary EVs have been also reported to be transferred from one organism to another and to influence mating behavior in Caenorhabditis elegans[63].
Similar to ciliogenesis, IS assembly and maintenance rely on the directional transport of endosomes containing receptors and signaling molecules that are known to exploit the recycling pathway to accumulate at the contact site with a cognate cell. Moreover, the IS serves as a focal point for both exocytosis and endocytosis that are triggered by TCR signaling at the plasma membrane[64]. The TCR itself constantly recycles from the cell surface to the recycling compartment and back to the plasma membrane as a mechanism of quality control of its signaling components[65][66]. However, upon engagement by a pMHC ligand, TCR surface levels are down-regulated as exhausted TCRs are internalized and sorted for recycling or lysosomal degradation[67]. Additionally, some of the post-endocytic TCRs together with a variety of cargoes, ranging from death receptor ligands, co-stimulatory receptors, microRNAs, genomic and mitochondrial DNA, are included in EVs that are focally released into the synaptic cleft, delivering signals to the target cell[68][69][70][71][72]. Polarized exocytosis is exploited also by activated T helper cells and CTLs for the focal release of vesicles containing cytokines or specialized secretory lysosomes carrying a battery of granzymes and the lytic pore-forming protein perforin[73][74].
4. Molecular similarities
In recent years many of the proteins labelled as “ciliary proteins”, as they have been discovered and then characterized in the context of the primary cilium, have been shown to also localize outside the primary cilium and to perform extraciliary functions, suggesting that these proteins play a general role in a range of different cellular processes (i.e. cell cycle, cytoskeleton re-organization, vesicular trafficking) and that are, directly or indirectly, linked to cell polarity.[75][76] Among others, ciliary proteins have been implicated in different mechanisms of IS assembly, ranging from centrosome polarization and docking at the plasma membrane, to actin cytoskeleton rearrangements and vesicular trafficking.
The ciliopathy-related Bardet-Biedl syndrome 1 component, a core component of the BBS complex that is mainly involved in the retrieval of membrane proteins (e.g. G-protein-coupled receptors) from the primary cilium into the cell[77][78], also participates in CD4+ T cell IS assembly by regulating the trafficking of proteasomal regulatory 19S subunit to the centrosome, where an increased activity of the local proteasome is required for centrosome polarization to the IS[41]. A peculiar feature of CTL IS is centrosome anchoring to the plasma membrane through the distal appendages of the mother centriole[11], allowing microtubules to re-organize just beneath the plasma membrane, thereby ensuring the directional transport of cytotoxic granules towards a specialized secretory domain within the synapse. The master distal appendage protein CEP83, which is required for ciliogenesis[79][80][81], shows a conserved function in non-ciliated CTLs, since its silencing by using RNA interference results in a reduced secretion of granules at the lytic synapse[11].
Consistent with the involvement of actin rearrangement in the assembly of both the primary cilium and the IS, the actin-related complex Arp2/3 is a shared participant in these processes. The Arp2/3 complex not only generates a branched actin network at the IS that facilitates T cell spreading[82][83] but also controls ciliogenesis and cilia length[29][31][39]. Furthermore, regulators of the tubulin cytoskeleton, such as the microtubule-associated protein 4 (MAP4) and the chaperonin containing T-complex protein (CCT, also known as TRiC) that controls cilia length and formation[84][85], have been later implicated in tubulin post-translational modifications and thus microtubule stability, on which centrosome polarization and vesicle dynamics depend[86][87].
Vesicular trafficking, and in particular the trafficking pathway responsible for TCR recycling to the IS, has been extensively investigated in recent years[88]. Surprisingly, some ciliogenesis-related proteins, which belong to the protein families of Rab GTPases together with their guanine nucleotide exchange factors (GEF)[89][90][91][92][93][94][95][96][97] and of soluble NSF attachment protein receptors (SNARE)[98], as well as some adaptor molecules[99][100][101][102] and IFT proteins[103], have now emerged as new players in IS assembly by controlling the synaptic recruitment of the TCR itself and of other receptors and signaling molecules that are known to accumulate at the IS[12][66][97][104][105][106][107][108][109][110][111][112][113] (e.g. the transferrin receptor, the chemokine receptor CXCR4, the transmembrane adaptor LAT, the kinase LCK). Most ciliary proteins specifically implicated in IS assembly have been mapped on the two main arms of the recycling pathway, regulated by such as Rab4 and Rab11, suggesting the existence of different subpopulations of recycling endosomes that, while participating in the canonical recycling pathways orchestrated by Rab4 and Rab11, are responsible for carrying specific cargo destined for the synaptic membrane.
The primary cilium is essential for the transduction of Hedgehog (Hh) signaling[114][115]. Binding of exogenous ligands (i.e. Sonic, Indian or Desert Hh) to the trans-membrane receptor Patched allows the ciliary translocation and activation of the signal transducer Smoothened and the subsequent activation of Gli and Gli-mediated transcription of target genes. Following TCR activation, CTLs express Hh pathway components, including Hh ligands, Patched, Smoothened and Gli. In the non-ciliated CTLs, Hh signaling promotes a Rac1-dependent remodeling of cortical actin, which is instrumental for centrosome polarization and CTL-mediated killing[116].
This entry is adapted from the peer-reviewed paper 10.3390/cells8080789