Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Subjects:
Biochemical Research Methods
The processing of berry juice, wine, or other beverages results in a considerable amount of pomace, including skins, seeds, and, occasionally, stalks. Pomace has been estimated at 30% of the total grape use in winemaking or 60% of the total cranberry use in juice production. However, berry pomace is no longer seen as a by-product and is further processed as a value-added food ingredient.
- berry pomace
- dietary fiber
- techno-functional properties
1. Introduction
The processing of berry juice, wine, or other beverages results in a considerable amount of pomace, including skins, seeds, and, occasionally, stalks. Pomace has been estimated at 30% of the total grape use in winemaking [1] or 60% of the total cranberry use in juice production [2]. However, berry pomace is no longer seen as a by-product and is further processed as a value-added food ingredient [3]. Multiple studies reported that antioxidant flavone fractions were extracted from berry pomace with high efficiency and evidenced strong free-radical-scavenging capacities [4][5][6]. Due to a high carbohydrate fraction, berry pomace can be considered a source of dietary fiber (DF).
The DF content in the pomace and the ratio between the soluble and insoluble fiber depend on the source and processing conditions during fiber isolation [7]. The pomace is typically deep-frozen at a minimum of −20 °C [8], milled, and dried at a temperature greater than 60 °C, resulting in a dry powder product. Reißner et al. [9] analyzed the chemical composition of dried black currant, red currant, gooseberry, rowanberry, and chokeberry pomaces. The authors reported that the insoluble fiber content in the dried pomaces varied between approximately 50.0 and 60.0 g/100 g DM and that the soluble fiber content varied between approximately 4.0 and 7.0 g/100 g DM. Other studies have highlighted the excellent technological properties (viscosity enhancement, swelling, water- and oil-holding capacities) of berry pomace [10]. Pomace from wine grapes [11] and pineapples [12] showed promising potential for use as an ingredient in the production of meat and dairy products to improve their stability and texture.
Moreover, the DF-rich berry pomace can promote health benefits, such as reducing preprandial cholesterol and postprandial blood glucose levels [13], enhancing gastrointestinal immunity [14], and increasing the satiety of consumers [15]. There are several pathways described regarding how DF influences glucose and cholesterol levels. For example, it increases the viscosity of the contents of the small intestine and retards the diffusion of glucose and lipids; it adsorbs glucose, cholesterol, and bile salts and decreases the levels of available glucose and lipids; it inhibits the activity of α-amylase and α-glucosidase; and it increases the formation of short-chain fatty acids in the gut and inhibits the formation of cholesterol [16][17].
Although the beneficial effects of DF have been well documented, their ability to deliver or promote these properties is highly dependent on their intake, as well as the source, physicochemical properties, and chemical composition. Thus, the characterization of berry pomace as a source of DF requires substantial knowledge regarding hypoglycemic activity and hypolipidemic effects. These properties of the berry pomace are particularly important when it is used as a food ingredient and the potential physiological effects must be predicted.
2. Chemical Composition and Functional Properties of the Berry PP
The proximate composition of the cranberry, lingonberry, sea buckthorn, and black currant PP is presented in Table 1. The PP of all four types of berries were composed mainly of dietary fiber, with smaller amounts of protein, fat, and ash. The protein content in the berry PP varied widely, from 21.09 ± 0.36 g/100 g DM for sea buckthorn PP to 7.4 ± 0.06 g/100 g DM for cranberry PP. The measured fat content was higher in the black currant PP (13.85 ± 0.27 g/100 g DM). The cranberry, lingonberry, and sea buckthorn PP contained 9.83 ± 0.46, 12.68 ± 0.39, and 12.95 ± 0.44 g/100 g DM of fat, respectively. The total dietary fiber (TDF) content ranged from 49.24 ± 0.95 g/100 g DM in the black currant PP to 73.85 ± 0.83 g/100 g DM in the lingonberry PP. The results of the analyses demonstrated the predominance of IDF in all the berry PP. Most of the IDF consisted of cellulose and acid-insoluble lignin. The cellulose content ranged from 7.45 ± 1.04 g/100 g DM in the black currant PP to 17.89 ± 1.88 g/100 g DM in the lingonberry PP. The acid-insoluble lignin content was approximately 39 g/100 g DM for the sea buckthorn and cranberry PP, 42 g/100 g DM for the lingonberry PP, and 30 g/100 g DM for the black currant PP. A higher ratio between the SDF and IDF was determined for the cranberry and black currant PP (0.21 and 0.20, respectively) and a lower ratio was observed for the lingonberry and sea buckthorn PP (0.13 and 0.08, respectively).
Table 1. Proximate chemical composition of berry pomace powder, g/100 g DM.
| Parameters | Cranberry | Lingonberry | Sea Buckthorn | Black Currant |
|---|---|---|---|---|
| Moisture | 5.57 c ± 0.11 | 3.41 a ± 0.04 | 4.17 b ± 0.04 | 7.97 d ± 0.10 |
| Ash | 0.96 a ± 0.04 | 1.18 b ± 0.01 | 1.38 c ± 0.04 | 3.82 d ± 0.02 |
| Protein (N × 6.25) | 7.4 a ± 0.06 | 8.60 b ± 0.27 | 21.09 c ± 0.36 | 9.05 b ± 0.26 |
| Fat | 9.83 a ± 0.46 | 12.68 b ± 0.39 | 12.57 b ± 0.20 | 13.85 c ± 0.27 |
| Total dietary fiber | 72.67 c ± 1.55 | 73.85 c ± 0.83 | 63.61 b ± 1.64 | 49.24 a ± 0.95 |
| Total insoluble dietary fiber: | 59.93 b ± 1.46 | 65.36 c ± 0.67 | 58.69 b ± 0.96 | 40.95 a ± 0.78 |
| Cellulose | 17.14 a ± 2.01 | 17.89 a ± 1.88 | 13.62 b ± 2.24 | 7.45 c ± 1.04 |
| Acid-insoluble lignin | 39.58 a ± 1.55 | 42.08 b ± 1.73 | 39.23 a ± 2.09 | 30.16 c ± 1.91 |
| Total soluble dietary fiber | 12.74 c ± 0.09 | 8.49 b ± 0.05 | 4.92 a ± 0.68 | 8.29 b ± 0.17 |
| Soluble DF/insoluble DF ratio | 0.21 a | 0.13 b | 0.08 c | 0.20 a |
| Carbohydrates * | 9.14 a | 4.43 b | 1.35 c | 24.04 d |
| Total phenolic content, GAE/g DM | 3.89 a ± 0.29 | 6.26 b ± 0.23 | 5.73 c ± 0.02 | 11.06 d ± 0.40 |
* Calculated as 100 − (fat + ash + protein + total dietary fiber). Different letters indicate statistically significant differences in row, p < 0.05.
Data on the chemical composition of berry pomace in the literature are scarce, and the reported values are not comparable due to the differences in pomace preparation and the analytical methods used. However, in the case of cranberry pomace, certain general and comparable compositional features have been established. Andreani and Karboune [18] reported a similar protein (7.6 g/100 g DM), fat (6.3 g/100 g DM), and ash (0.9 g/100 g DM) content and a slightly lower value for the total dietary fiber (63 g/100 g DM) for dried cranberry pomace. Data in the literature on the basic components of dried black currant pomace vary widely, for example, from 11.8 to 15.7 g/100 g DM of protein, 2.5 to 20.2 g/100 g DM of fat, 2.3 to 4.1 g/100 g DM of ash, and 59.1 to 67.4 g/100 g DM of total dietary fiber [9][19][20]. This high variability may be due to differences in the cultivation, ripeness, and processing conditions of the berry pomace. Regarding the chemical composition of the sea buckthorn pomace, obtained results for the protein and ash content were similar to those reported by Ben-Mahmoud et al. [21] for dried sea buckthorn pomace (20.87 and 2.02%, respectively). The fat content observed in the present study was similar to that reported in studies performed by Ben-Mahmoud et al. [21] and Nour et al. [22]. The authors reported that the sea buckthorn pomace that was used as a supplement in animal feed consisted of 10.52 to 20.05% fat. However, the data in the literature on fiber content only include the crude fiber content in dried sea buckthorn pomace, which ranges from 18.3 to 19.86% [21][22]. According to Hao et al. [23], sea buckthorn pomace contains 29.27 to 32.11% DM of neutral detergent fiber and 20.45 to 22.9% DM of acid detergent fiber.
Most of the berry sugars and phenolic compounds were extracted into the juice. However, the residual sugar content in the berry PP was characterized by a great variability and ranged from 1.77 g/100 g DM for sea buckthorn PP to 24.04 g/100 g DM for black currant PP. The total phenolic content in the PP was also dependent on the type of berry and varied from 3.89 to 11.06 GAE/g DM. This content was higher than the total phenol content observed in seedless black currant pomace [20] and was similar to the results reported for blueberry and cranberry pomaces [24]. The fact that black currant PP contained the highest quantity of sugars and phenolic compounds indicates that the effectiveness of the juice extraction from the black currants was low.
The chemical composition of berry PP, particularly the amount and proportion of the SDF and IDF, leads to great variation in their functionality (Table 2).
Table 2. Physicochemical properties of berry pomace powder.
| Powder of Pomace | Water Holding Capacity, (g/g) | Swelling Capacity, (mL/g) | Oil Binding Capacity, (g/g) | ||
|---|---|---|---|---|---|
| pH 2 | pH 7 | pH 2 | pH 7 | ||
| Black currant | 2.78 a ± 0.04 | 2.78 a ± 0.02 | 4.14 a ± 0.25 | 4.99 a ± 0.21 | 1.14 a ± 0.01 |
| Cranberry | 3.83 b ± 0.09 | 3.87 b ± 0.18 | 7.95 b ± 0.40 | 7.99 b ± 0.40 | 1.57 b ± 0.05 |
| Lingonberry | 3.28 c ± 0.05 | 3.27 c ± 0.02 | 7.90 b ± 0.40 | 8.00 b ± 0.40 | 1.46 c ± 0.05 |
| Sea buckthorn | 4.32 d ± 0.04 | 4.24 d ± 0.15 | 10.95 c ± 0.50 | 9.98 c ± 0.55 | 1.09 d ± 0.02 |
Different letters indicate statistically significant differences in column, p < 0.05.
The physicochemical properties of the berry PP, such as the WHC, SC, and OBC, are important from a technological and physiological standpoint. They are essential for the successful introduction of fiber-rich PP as a food ingredient and contribute to the textural properties and stability of formulated foods [3]. Additionally, the physicochemical properties of fiber-rich food ingredients are relevant for their hypoglycemic activity and hypolipidemic effects [25].
The WHC and SC of the berry PP at pH 7 ranged from 2.78 to 4.24 g/g and 4.99 to 9.98 mL/g, respectively. The highest WHC and SC were recorded for sea buckthorn PP. Similar values were observed for the WHC and SC when measured at pH 2. Previous studies on the hydration properties of red raspberry pomace reported similar results (WHC of 2.62 g/g and SC of 6.93 mL/g) [26]. A study conducted on pear pomace also exhibited similar WHC and SC values (3.44 g/g and 5.09 mL/g, respectively) [27]. Gouw et al. [24] reported higher WHC and lower SC values for dried berry pomace (a WHC of 8.70 g/g for cranberry pomace and 7.71 g/g for red raspberry pomace, and an SC of 5.87 mL/g for cranberry pomace and 2.88 mL/g for red raspberry pomace). The OHC of the berry PP in the present study ranged from 1.09 to 1.57 g/g and was highest for the cranberry PP, while the sea buckthorn PP showed the lowest value regarding the OHC. A high SDF content in cranberry PP may contribute to an increase in the OHC [28]. Previous studies reported higher OHC values for red raspberry pomace (2.44 g/g, [26]), cranberry and blueberry pomace (1.97 and 1.96 g/g, respectively, [24]), and pear pomace (1.82 g/g, [27]) and considerably higher OHC values for orange and grapefruit DF (3.62 and 8.20 g/g, respectively, [29]).
When berry PP is observed to have good hydration properties, the potential for its use as a food ingredient with a certain functionality can be predicted. This functionality may be reflected in certain physiological effects, such as hypoglycemic and hypolipidemic effects.
3. In Vitro Hypoglycemic Effects
The GAC is an index that characterizes the behavior of fiber in glucose adsorption during gastrointestinal transit in vitro [30]. The GAC is associated with the physical properties of the network structure of DF, which may cause the adsorption or inclusion of small sugar molecules. Therefore, the consumption of DF leads to the inhibition of postprandial hyperglycemia [29]. The GAC of the berry PP tested in the present study is presented in Figure 1. The GAC was dependent on the molar concentration of glucose. At low glucose concentrations (5 and 10 mmol/L), all the berry PP exhibited a small and similar capacity to adsorb glucose, with values ranging from 0.05 to 0.12 mmol/g. As the concentration of glucose increased to 100 mmol/L, the berry PP effectively increased the GAC, with the black currant and sea buckthorn PP showing the highest values (1.20 and 1.31 mmol/g, respectively). These results indicated that the berry PP were effective at adsorbing glucose only at high concentrations (50 and 100 mmol/L), although the quantity of glucose bound to the berry PP was dependent on the source of the PP. Pomaces from various fruit and vegetable sources have been reported to exhibit a GAC, most likely due to a high IDF content. For example, the GAC of DF extracted from carrots was shown to be dependent on the extraction method used and ranged from 2.43 to 2.63 mmol/g after incubation in a 100 mmol/L glucose solution [31]. In a separate report, de-oiled red raspberry pomace and DF extracted from de-oiled red raspberry pomace showed GAC values of 3.42 mg/g and 1.73 mg/g, respectively [26]. Huang et al. [32] observed a GAC value of 0.89 mmol/g for psyllium pomace after incubation in a 200 mmol/L glucose solution. The authors also demonstrated that, with increased glucose concentrations, higher amounts of glucose were bound. The results of the present study suggest that the berry PP can maintain a low level of glucose in the small intestine, which reduces the contact with the intestinal tract and, consequently, decreases the potential for postprandial hyperglycemia.
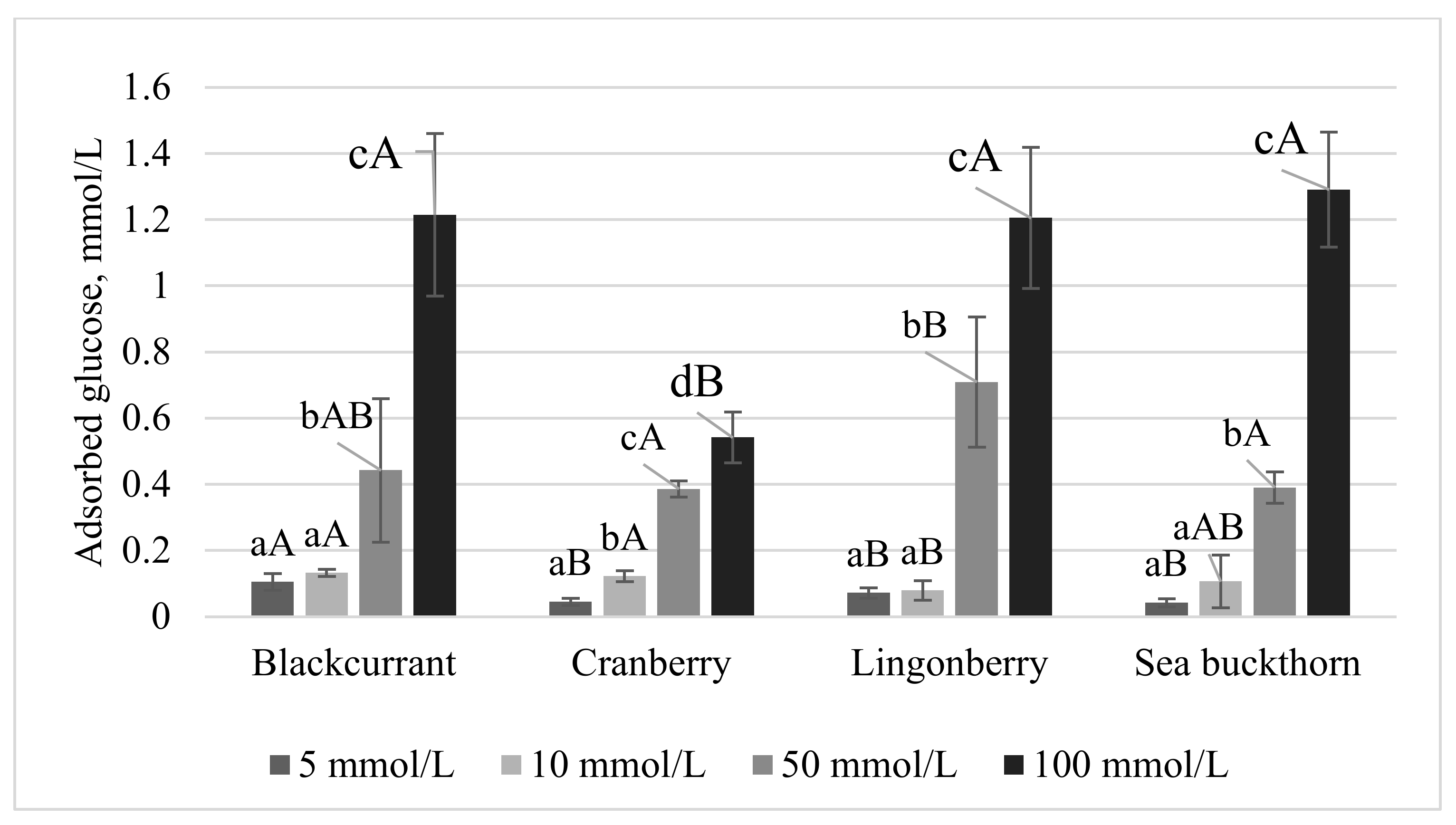 Figure 1. Glucose adsorption capacity at different concentrations of glucose (5, 10, 50, and 100 mmol/L) of berry PP. Values expressed as average ± st. dev. Different letters among columns indicate significant (p < 0.05) differences (lower case—among concentrations; capital letters—among berry PP with the same glucose concentration).
Figure 1. Glucose adsorption capacity at different concentrations of glucose (5, 10, 50, and 100 mmol/L) of berry PP. Values expressed as average ± st. dev. Different letters among columns indicate significant (p < 0.05) differences (lower case—among concentrations; capital letters—among berry PP with the same glucose concentration).The GDRI is an index that is used to characterize the hypoglycemic effects of DF. The GDRI was created to predict the effect of fiber on delaying glucose absorption in the gastrointestinal tract due to glucose entrapment by the network formed by DF [33]. The in vitro glucose diffusion and GDRI of the berry PP tested in the present study are presented in Table 3.
Table 3. Effect of berry PP on glucose diffusion from 30 to 180 min of incubation at 37 °C.
| Powder of Pomace | Glucose Concentration in the Dialysate, (mmol/g) | |||
|---|---|---|---|---|
| 30 min | 60 min | 120 min | 180 min | |
| Control | 1.15 aA ± 0.01 | 1.58 bA ± 0.02 | 1.98 cA ± 0.03 | 2.10 cA ± 0.1 |
| Black currant | 0.96 aB ± 0.05 (16.61) * | 1.40 bB ± 0.07 (11.39) | 1.77 cB± 0.09 (10.96) | 1.92 cB ± 0.10 (8.46) |
| Cranberry | 1.03 aB ± 0.05 (10.40) | 1.48 bB ± 0.07 (6.40) | 1.86 cB ± 0.09 (5.97) | 2.05 cA ± 0.10 (2.22) |
| Lingonberry | 0.83 aC ± 0.04 (27.49) | 1.17 bC ± 0.06 (25.74) | 1.62 cC ± 0.08 (18.50) | 1.83 cB ± 0.09 (12.76) |
| Sea buckthorn | 1.00 aB ± 0.05 (13.30) | 1.48 bB ± 0.07 (6.13) | 1.93 cAB ± 0.10 (2.49) | 2.05 cA ± 0.10 (2.23) |
* Data in parentheses present the glucose dialysis retardation indexes of various samples. Different letters among columns indicate significant (p < 0.05) differences (lower case—among time of dialysis; capital letters—among berry PP at the same time of dialysis).
During a time interval from 0 to 180 min, the glucose concentration increased in the dialysate for all the samples. In the case of the sea buckthorn PP, the registered increase in released glucose ranged from 1.00 mmol (after 30 min) to 2.05 mmol (after 180 min). For the lingonberry PP, the glucose concentration in the dialysate increased from 0.83 mmol (after 30 min) to 1.83 mmol (after 180 min). However, the control sample (that did not contain the berry PP) displayed a significantly higher glucose release (p < 0.05) at various time intervals compared to the berry PP samples. By calculating the GDRI, which indicates the effect of the berry PP on the delay in glucose absorption in the gastrointestinal tract, it was obvious that all the berry PP decreased the amount of glucose dialyzed regarding the control sample. After 30 min, the lingonberry PP exhibited the highest GDRI (27.49%), followed by the black currant PP (16.61%), the sea buckthorn PP (13.30%), and the cranberry PP (10.40%). Similar values were observed after 60, 130, and 180 min of dialysis. Acquired findings are in accordance with the observations reported for DF isolated from orange and psyllium pomace [32] and DF-rich coffee parchment [34]. However, the values observed in the present study were lower than those reported for other sources of DF, such as oats, barley, and psyllium husk [35]. This capability of DF to delay glucose absorption is associated with the increased viscosity of soluble polysaccharides and the network formed by the insoluble fiber that maintains the entrapped glucose [36]. In the present study, we assumed that the latter prevailed regarding the decrease in glucose absorption. According to our results, the lingonberry PP, which showed the highest amount of IDF and a moderate amount of SDF (65.36 and 8.49%, respectively), exhibited the highest GDRI values. Moreover, the cranberry PP, which showed a lower IDF content and a high SDF content (59.93 and 12.74%, respectively), exhibited the lowest GDRI.
4. In Vitro Hypolipidemic Effects
The CBC and NaChBC are valuable indicators for characterizing the hypolipidemic properties of fiber-rich berry PP [37]. The results recorded in the present study regarding these capacities are presented in Table 4.
Table 4. Cholesterol- and sodium-cholate-binding capacities of berry pomace powder.
| Powder of Pomace | Cholesterol-Binding Capacity, mg/g | Sodium-Cholate-Binding Capacity, mg/g | |
|---|---|---|---|
| pH 2 | pH 7 | pH 7 | |
| Black currant | 18.02 a ± 0.01 | 21.11 a ± 0.42 | 74.78 a ± 1.39 |
| Cranberry | 21.91 b ± 0.02 | 23.13 b ± 0.47 | 52.68 b ± 2.07 |
| Lingonberry | 14.16 c ± 0.01 | 22.61 b ± 0.45 | 40.71 c ± 2.78 |
| Sea buckthorn | 15.11 d ± 0.06 | 22.75 b ± 0.46 | 24.66 d ± 5.80 |
Different letters among columns indicate significant differences (p < 0.05).
The CBC of the cranberry, lingonberry, sea buckthorn, and black currant PP at pH 7 were 23.13 ± 0.47 mg/g, 22.61 ± 0.45 mg/g, 22.75 ± 0.46 mg/g, and 21.11 ± 0.42 mg/g, respectively. These results were in line with the OBC of tested berry PP (Table 4)—the highest OBC was observed for the cranberry PP and the lowest OBC was observed for the sea buckthorn and black currant PP. At pH 2, all the berry PP tested bind a lower amount of cholesterol. These results corroborate those obtained by other researchers who observed that cholesterol adsorption by DF occurred mainly in the intestines rather than the stomach [38]. The cranberry PP demonstrated the highest CBC (21.91 ± 0.02 mg/g) and the lingonberry PP showed the lowest CBC at pH 2. A decrease in the cholesterol levels in the small intestine is associated with SDF rather than IDF [39]. In the present study, the cranberry PP contained the highest amount of SDF (12.74 ± 0.09 mg/100 g DM), which may be associated with the improved CBC observed for this PP compared with the others. There is an abundance of scientific evidence regarding the antihyperlipidemic activity of polyphenol-rich berries on the key steps of lipid digestion and absorption [25]. However, data on the capacity of the DF from berries to bind cholesterol are not so abundant. In a previous study, the cholesterol adsorption capacity and the total DF for de-oiled red raspberry pomace were reported at 6.04 mg/g and 2.61 mg/g (at pH 7), respectively [26]. Additionally, data exists on the hypolipidemic effects of DF from various fruits. Dried fiber-rich orange pomace contained 17.31% of SDF and showed 6.89 mg/g DM of CBC [32]. In the case of dried peach pomace that contained 10.0 g/100 g of SDF, the recorded CBC was 3.88 ± 0.48 mg/g at pH 7 and 1.27 ± 0.02 mg/g at pH 2 [27]. Wang et al. [29] reported that the CBC of SDF from various citrus fruit peels at pH 2 ranged from 1.66 ± 0.29 mg/g to 10.88 ± 0.32 mg/g, while the values were recorded at 3.22 ± 0.38 mg/g to 17.90 ± 0.17 mg/g at pH 7. Moreover, the CBC of SDF from wheat bran was considerably lower at 2.17 ± 0.07 mg/g and 3.48 ± 0.03 mg/g at pH 2 and pH 7, respectively [40].
DF is also capable of trapping bile acids in the small intestine. In the human body, more than 90% of bile acids exist in the form of binding substances, such as sodium cholate. Obtained results demonstrated that the binding capacity of sodium cholate for four DF-rich berry PP were different. The highest NaChBC was observed for the black currant PP (74.78 ± 1.39 mg/g) at pH 7, followed by the cranberry, lingonberry, and sea buckthorn PP. These bile-acid-binding values are comparable to those reported for modified millet bran DF [41] and are higher than those reported for DF in soybean-seed hulls, wheat bran, and apple peels [40], but they are considerably lower compared to the SDF recorded in citrus fruits peels [29].
Various mechanisms have been described regarding the reduction of the NaChBC by fiber. The main effects are associated with the viscous and gel-forming properties of soluble fiber and the presence of phenolic compounds. The presence of two types of binding sites on the bile salt micelles makes them suitable for binding both hydrophobic and hydrophilic bioactive molecules [42]. Zhou and Wang [43] revealed that phenolic compounds may bind with sodium cholate monomers, dimers, and primary and secondary micelles mainly through hydrophobic interactions. In the present study, the black currant PP was high in phenolic compounds (11.06 ± 0.40 GAE/g DM), and this can impact the high capacity of this PP to bind sodium cholate. The cranberry PP also showed a high sodium-cholate-binding capacity and contained a higher amount of SDF (12.74 ± 0.09 mg/100 g DM) compared to the other berry PP. According to Ma et al. [44], the effectiveness of DF in increasing sodium cholate binding is mainly attributed to a high water SC and SDF content, which can increase the viscosity of the medium.
Thus, the berry PP can effectively absorb bile acids and other lipid substances in the small intestine and help to remove these substances while performing as a lipid scavenger [45]. A decrease in sodium cholate content invokes the conversion of cholesterol into sodium cholate for its supplementation, thereby promoting the consumption of cholesterol [46]. Therefore, the adsorption capacity of DF for cholesterol and cholate is a good indicator for the adsorption of lipophilic substances. The differences in the hypolipidemic effects of the berry PP tested in the present study were associated with their composition.
This entry is adapted from the peer-reviewed paper 10.3390/foods11050716
References
- De Camargo, A.C.; Regitano-d’Arce, M.A.B.; Biasoto, A.C.T.; Shahidi, F. Low molecular weight phenolics of grape juice and winemaking byproducts: Antioxidant activities and inhibition of oxidation of human low-density lipoprotein cholesterol and dna strand breakage. J. Agric. Food Chem. 2014, 62, 12159–12171.
- Tokusoglu, Ö.; Hall, C.A., III. Fruit and Cereal Bioactives, 1st ed.; CRC Press: Boca Raton, FL, USA, 2011; pp. 3–8.
- Rohm, H.; Brennan, C.; Turner, C.; Günther, E.; Campbell, G.; Hernando, I.; Struck, S.; Kontogiorgos, V. Adding value to fruit processing waste: Innovative ways to incorporate fibers from berry pomace in baked and extruded cereal-based foods—A SUSFOOD Project. Foods 2015, 4, 690–697.
- Harrison, J.E.; Oomah, B.D.; Diarra, M.S.; Ibarra-Alvarado, C. Bioactivactivities of pilot-scale extracted cranberry juice and pomace. J. Food Process. Pres. 2013, 37, 356–365.
- Ravi, H.K.; Breil, C.; Vian, M.A.; Chemat, F.; Venskutonis, P.R. Biorefining of Bilberry (Vaccinium Myrtillus L.) pomace using microwave hydrodiffusion and gravity, ultrasound-assisted, and bead-milling extraction. ACS Sustain. Chem. Eng. 2018, 6, 4185–4193.
- Tamkutė, L.; Liepuoniūtė, R.; Pukalskienė, M.; Venskutonis, P.R. Recovery of valuable lipophilic and polyphenolic fractions from cranberry pomace by consecutive supercritical CO2 and pressurized liquid extraction. J. Supercrit. Fluids 2020, 159, 104755.
- Tanongkankit, Y.; Sablani, S.S.; Chiewchan, N.; Devahastin, S. Microwave-assisted extraction of sulforaphane from white cabbages: Effects of extraction condition, solvent and sample pretreatment. J. Food Eng. 2013, 117, 151–157.
- Struck, S.; Plaza, M.; Turner, C.; Rohm, H. Berry pomace—A review of processing and chemical analysis of its polyphenols. Int. J. Food Sci. Technol. 2016, 51, 1305–1318.
- Reißner, A.-M.; Al-Hamimi, S.; Quiles, A.; Schmidt, C.; Struck, S.; Hernando, I.; Turner, C.; Rohm, H. Composition and physicochemical properties of dried berry pomace: Composition and technofunctional properties of berry pomace. J. Sci. Food Agric. 2019, 99, 1284–1293.
- Alba, K.; Campbell, G.M.; Kontogiorgos, V. Dietary fibre from berry processing waste and its impact on bread structure: A review. J. Sci. Food Agric. 2019, 99, 4189–4199.
- Tseng, A.; Zhao, Y. Wine grape pomace as antioxidant dietary fibre for enhancing nutritional value and improving storability of yogurt and salad dressing. Food Chem. 2013, 138, 356–365.
- Sah, B.N.P.; Vasiljevic, T.; McKechnie, S.; Donkor, O.N. Physicochemical, textural and rheological properties of probiotic yogurt fortified with fibre-rich pineapple peel powder during refrigerated storage. LWT Food Sci. Technol. 2016, 65, 978–986.
- Hauner, H.; Bechthold, A.; Boeing, H.; Brönstrup, A.; Buyken, A.; Leschik-Bonnet, E.; Linseisen, J.; Schulze, M.; Strohm, D.; Wolfram, G. Evidence-based guideline of the german nutrition society: Carbohydrate intake and prevention of nutrition-related diseases. Ann. Nutr. Metab. 2012, 60, 1–58.
- Gunness, P.; Gidley, M.J. Mechanisms underlying the cholesterol-lowering properties of soluble dietary fibre polysaccharides. Food Funct. 2010, 1, 149.
- Brownlee, I.A. The physiological roles of dietary fibre. Food Hydrocoll. 2011, 25, 238–250.
- Ou, S.; Kwok, K.; Li, Y.; Fu, L. In vitro study of possible role of dietary fiber in lowering postprandial serum glucose. J. Agric. Food Chem. 2001, 49, 1026–1029.
- Kabir, A.U.; Samad, M.B.; D’Costa, N.M.; Akhter, F.; Ahmed, A.; Hannan, J. Anti-HYPERGLYCEMIC Activity of Centella Asiatica Is Partly Mediated by Carbohydrase Inhibition and Glucose-Fiber Binding. BMC Complement. Altern. Med. 2014, 14, 31.
- Spadoni Andreani, E.; Karboune, S. Comparison of Enzymatic and Microwave Assisted Alkaline Extraction Approaches for the Generation of Oligosaccharides from American Cranberry (Vaccinium macrocarpon) Pomace. J. Food Sci. 2020, 85, 2443–2451.
- Alberici, N.; Fiorentini, C.; House, A.; Dordoni, R.; Bassani, A.; Spigno, G. Enzymatic Pre-Treatment of Fruit Pomace for Fibre Hydrolysis and Antioxidants Release. Chem. Eng. Trans. 2020, 79, 175–180.
- Sójka, M.; Król, B. Composition of Industrial Seedless Black Currant Pomace. Eur. Food Res. Technol. 2009, 228, 597–605.
- Ben-Mahmoud, Z.; Mohamed, M.S.; Bláha, J.; Lukešová, D.; Kunc, P. The Effect of Sea Buckthorn (Hippophae rhamnoides L.) Residues in Compound Feeds on the Performance and Skin Color of Broilers. Int. J. Anim. Res. 2014, 48, 548.
- Nour, V.; Panaite, T.D.; Corbu, A.R.; Ropota, M.; Turcu, R.P. Nutritional and Bioactive Compounds in Dried Sea-Buckthorn Pomace. Erwerbs-Obstbau 2021, 63, 91–98.
- Hao, X.Y.; Ding, N.; Mu, C.T.; Zhang, C.X.; Zhao, J.X.; Zhang, J.X. Effects of Sea Buckthorn Pomace Supplementation on Energy Partitioning and Substrate Oxidation in Male Lambs. Anim. Feed Sci. Technol. 2019, 247, 149–156.
- Gouw, V.P.; Jung, J.; Zhao, Y. Functional properties, bioactive compounds, and in vitro gastrointestinal digestion study of dried fruit pomace powders as functional food ingredients. LWT Food Sci. Technol. 2017, 80, 136–144.
- Jiang, H.; Zhang, W.; Li, X.; Xu, Y.; Cao, J.; Jiang, W. The anti-obesogenic effects of dietary berry fruits: A review. Food Res. Int. 2021, 147, 110539.
- Li, M.; Liu, Y.; Yang, G.; Sun, L.; Song, X.; Chen, Q.; Bao, Y.; Luo, T.; Wang, J. Microstructure, physicochemical properties, and adsorption capacity of deoiled red raspberry pomace and its total dietary fiber. LWT Food Sci. Technol. 2022, 153, 112478.
- Yan, L.; Li, T.; Liu, C.; Zheng, L. Effects of high hydrostatic pressure and superfine grinding treatment on physicochemical/functional properties of pear pomace and chemical composition of its soluble dietary fibre. LWT Food Sci. Technol. 2019, 107, 171–177.
- Chu, J.; Zhao, H.; Lu, Z.; Lu, F.; Bie, X.; Zhang, C. Improved physicochemical and functional properties of dietary fiber from millet bran fermented by bacillus natto. Food Chem. 2019, 294, 79–86.
- Wang, L.; Xu, H.; Yuan, F.; Pan, Q.; Fan, R.; Gao, Y. Physicochemical characterization of five types of citrus dietary fibers. Biocatal. Agric. Biotechnol. 2015, 4, 250–258.
- Peerajit, P.; Chiewchan, N.; Devahastin, S. Effects of pretreatment methods on health-related functional properties of high dietary fibre powder from lime residues. Food Chem. 2012, 132, 1891–1898.
- Yu, G.; Bei, J.; Zhao, J.; Li, Q.; Cheng, C. Modification of Carrot (Daucus carota Linn. Var. Sativa Hoffm.) pomace insoluble dietary fiber with complex enzyme method, ultrafine comminution, and high hydrostatic pressure. Food Chem. 2018, 257, 333–340.
- Huang, Y.-L.; Ma, Y.-S.; Tsai, Y.-H.; Chang, S.K.C. In vitro hypoglycemic, cholesterol-lowering and fermentation capacities of fiber-rich orange pomace as affected by extrusion. Int. J. Biol. Macromol. 2019, 124, 796–801.
- Nsor-Atindana, J.; Zhong, F.; Mothibe, K.J. In vitro hypoglycemic and cholesterol lowering effects of dietary fiber prepared from Cocoa (Theobroma cacao L.) Shells. Food Funct. 2012, 3, 1044.
- Benitez, V.; Rebollo-Hernanz, M.; Hernanz, S.; Chantres, S.; Aguilera, Y.; Martin-Cabrejas, M.A. Coffee parchment as a new dietary fiber ingredient: Functional and physiological characterization. Food Res. Int. 2019, 122, 105–113.
- Ahmed, F.; Sairam, S.; Urooj, A. In vitro hypoglycemic effects of selected dietary fiber sources. J. Food Sci. Technol. 2011, 48, 285–289.
- Céspedes, M.A.L.; Martínez Bustos, F.; Kil chang, Y. The effect of extruded orange pulp on enzymatic hydrolysis of starch and glucose retardation index. Food Bioproc. Technol. 2010, 3, 684–692.
- Luo, X.; Wang, Q.; Zheng, B.; Lin, L.; Chen, B.; Zheng, Y.; Xiao, J. Hydration properties and binding capacities of dietary fibers from bamboo shoot shell and its hypolipidemic effects in mice. Food Chem. Toxicol. 2017, 109, 1003–1009.
- Zhu, Y.; He, C.; Fan, H.; Lu, Z.; Lu, F.; Zhao, H. Modification of foxtail millet (Setaria Italica) bran dietary fiber by xylanase-catalyzed hydrolysis improves its cholesterol-binding capacity. LWT Food Sci. Technol. 2019, 101, 463–468.
- Rodríguez, R.; Jiménez, A.; Fernández-Bolaños, J.; Guillén, R.; Heredia, A. Dietary fibre from vegetable products as source of functional ingredients. Trends Food Sci. Technol. 2006, 17, 3–15.
- Zhang, N.; Huang, C.; Ou, S. In vitro binding capacities of three dietary fibers and their mixture for four toxic elements, cholesterol, and bile acid. J. Hazard. Mater. 2011, 186, 236–239.
- Zheng, Y.; Wang, X.; Tian, H.; Li, Y.; Shi, P.; Guo, W.; Zhu, Q. Effect of four modification methods on adsorption capacities and in vitro hypoglycemic properties of millet bran dietary fibre. Food Res. Int. 2021, 147, 110565.
- O’Connor, C.J.; Wallace, R.G. Physico-chemical behavior of bile salts. Adv. Colloid Interface Sci. 1985, 22, 1–111.
- Zhou, H.; Wang, X. Spectrometric study on the interaction of sodium cholate aggregates with quercetin. Colloids Surf. A Physicochem. Eng. Asp. 2015, 481, 31–37.
- Ma, M.; Mu, T. Effects of extraction methods and particle size distribution on the structural, physicochemical, and functional properties of dietary fiber from deoiled cumin. Food Chem. 2016, 194, 237–246.
- Devi, P.B.; Vijayabharathi, R.; Sathyabama, S.; Malleshi, N.G.; Priyadarisini, V.B. Health benefits of finger millet (Eleusine Coracana L.) polyphenols and dietary fiber: A review. J. Food Sci. Technol. 2014, 51, 1021–1040.
- Zhu, Y.; Chu, J.; Lu, Z.; Lv, F.; Bie, X.; Zhang, C.; Zhao, H. Physicochemical and functional properties of dietary fiber from foxtail millet (Setaria Italic) bran. J. Cereal Sci. 2018, 79, 456–461.
This entry is offline, you can click here to edit this entry!