Malaria is a vector-transmitted parasite disease that continues to plague mankind. It is caused in humans by five main species of Plasmodium. The majority of conventional antimalarials kill parasites via direct or indirect overproduction of reactive oxygen species (ROS). Even when some parasites try to manage these ROS, over production of the ROS still leads to parasite death. This therefore underscores the role of ROS in the antiplasmodial activity of different antimalarials.
- ROS
- antimalarials
- Plasmodium falciparum
- malaria and oxidative stress
1. ROS in Living Cells
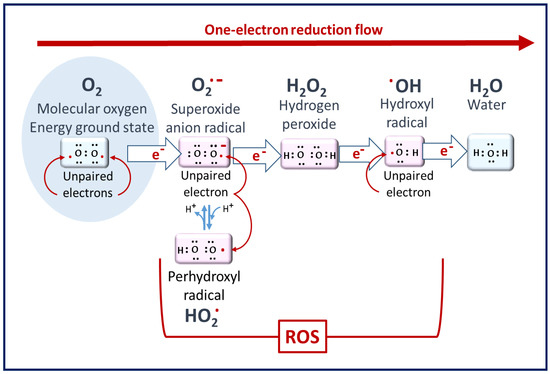
2. The Biochemical Impacts of ROS in Living Cells
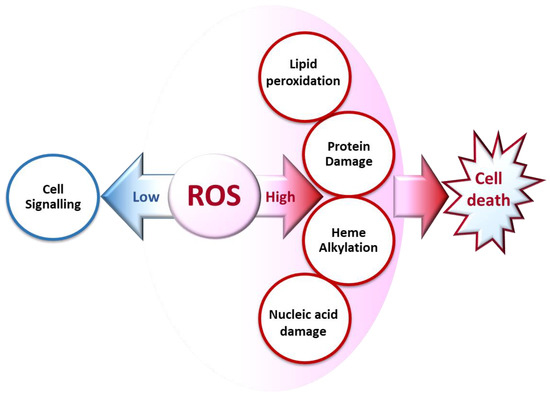
2.1. Cell Signaling
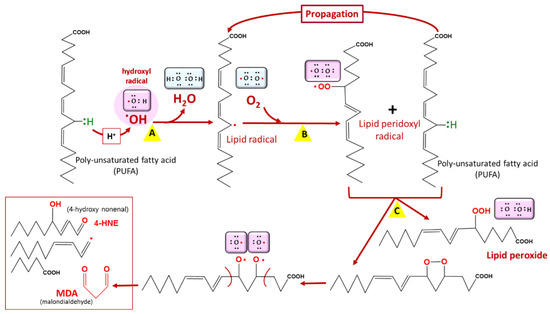
2.2. Lipid Peroxidation
2.3. Protein Damage
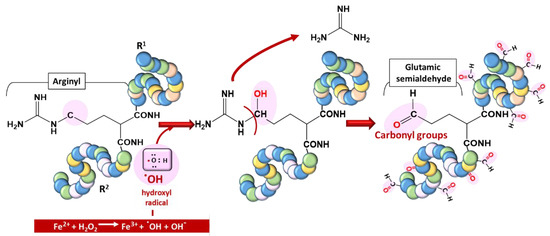
(ii) by secondary reactions on lysine, cysteine, and histidine with reactive carbonyl derivatives, resulting, among other things, from lipid peroxidation.
2.4. Nucleic Acid Damage
3. ROS in Plasmodium-Infected Erythrocytes under Steady State
| Agent | Site of Production | Role in Oxidative Homeostasis |
|---|---|---|
| HRP | FV | Binding with heme for polymerization |
| HDP | FV | Heme polymerization to hemozoin |
| H2O2 | Cytosol and FV | Degrades heme |
| SOD | Cytosol, Mitochondria | Dismutation of O2•− |
| Prx | Cytosol, Mitochondria, Apicoplast | Reduction of H2O2 to H2O |
| Trx | Cytosol, Mitochondria | Reduction of Prx |
| GSH | Cytosol | Degradation of heme, reduction of proteins and ROS |
| Vit B6 | Cytosol | Role unclear |
4. ROS Production in Plasmodium-Infected Erythrocytes under Antimalarial Treatment
4.1. Mode of Action of Chloroquine and Other Quinolines
4.2. Mode of Action of Atovaquone and Hydroxynaphtoquinones
4.3. Mode of Action of Artemisinin: The Role of Endoperoxide
4.3.1. Activation of the Endoperoxide to Generate ROS
4.3.2. Depolarization of the Mitochondrial Membrane Potential
4.4. Artemisinin-Based Combination Therapies
4.5. ROS Evasive Mechanisms under Treatment
4.5.1. Preventive Mechanisms
4.5.2. Reductive Mechanisms
4.5.3. Reparative Mechanisms
This entry is adapted from the peer-reviewed paper 10.3390/antiox10121872
References
- Santo, A.; Zhu, H.; Li, Y.R. Free Radicals: From Health to Disease. React. Oxyg. Species 2016, 2, 245–263.
- Li, Y.R.; Trush, M. Defining ROS in Biology and Medicine. React. Oxyg. Species 2016, 1, 9–21.
- Hancock, J.T.; Desikan, R.; Neill, S.J. Role of reactive oxygen species in cell signalling pathways. Biochem. Soc. Trans. 2001, 2, 345–350.
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive Oxygen Species in Inflammation and Tissue Injury. Antioxid. Redox Signal. 2014, 20, 1126.
- Stefanatos, R.; Sanz, A. The role of mitochondrial ROS in the aging brain. FEBS Lett. 2018, 592, 743–758.
- Sugamura, K.; Keaney, J.F. Reactive oxygen species in cardiovascular disease. Free Radic. Biol. Med. 2011, 51, 978–992.
- Deavall, D.G.; Martin, E.A.; Horner, J.M.; Roberts, R. Drug-induced oxidative stress and toxicity. J. Toxicol. 2012, 2012, 645460.
- Thannickal, V.J.; Fanburg, B.L. Reactive oxygen species in cell signaling. Am. J. Physiol. Lung Cell. Mol. Physiol. 2000, 279, L1005–L1028.
- Sampson, C.; Keens, R.H.; Kattnig, D.R. On the magnetosensitivity of lipid peroxidation: Two- versus three-radical dynamics. Phys. Chem. Chem. Phys. 2019, 21, 13526–13538.
- Hartwig, C.L.; Rosenthal, A.S.; D’Angelo, J.; Griffin, C.E.; Posner, G.H.; Cooper, R.A. Accumulation of artemisinin trioxane derivatives within neutral lipids of Plasmodium falciparum malaria parasites is endoperoxide-dependent. Biochem. Pharmacol. 2009, 77, 322–336.
- Kumura, N.; Furukawa, H.; Onyango, A.N.; Izumi, M.; Nakajima, S.; Ito, H.; Hatano, T.; Kim, H.S.; Wataya, Y.; Baba, N. Different behavior of artemisinin and tetraoxane in the oxidative degradation of phospholipid. Chem. Phys. Lipids 2009, 160, 114–120.
- Fernando, N.; Wickremesinghe, S.; Niloofa, R.; Rodrigo, C.; Karunanayake, L.; De Silva, H.J.; Wickremesinghe, A.R.; Premawansa, S.; Rajapakse, S.; Handunnetti, S.M. Protein carbonyl as a biomarker of oxidative stress in severe leptospirosis, and its usefulness in differentiating leptospirosis from dengue infections. PLoS ONE 2016, 11, e0156085.
- Dalle-Donne, I.; Rossi, R.; Giustarini, D.; Milzani, A.; Colombo, R. Protein carbonyl groups as biomarkers of oxidative stress. Clin. Chim. Acta 2003, 329, 23–38.
- Nyström, T. Role of oxidative carbonylation in protein quality control and senescence. EMBO J. 2005, 24, 1311–1317.
- Fedorova, M.; Bollineni, R.C.; Hoffmann, R. Protein carbonylation as a major hallmark of oxidative damage. Mass Spectrom Rev 2014, 33, 79–97.
- Thomas, J.A.; Tan, M.S.Y.; Bisson, C.; Borg, A.; Umrekar, T.R.; Hackett, F.; Hale, V.L.; Vizcay-Barrena, G.; Fleck, R.A.; Snijders, A.P.; et al. A protease cascade regulates release of the human malaria parasite Plasmodium falciparum from host red blood cells. Nat. Microbiol. 2018, 3, 447–455.
- Rosenthal, P.J. Cysteine proteases of malaria parasites. Int. J. Parasitol. 2004, 34, 1489–1499.
- Wu, W.M.; Chen, Y.L.; Zhai, Z.; Xiao, S.H.; Wu, Y.L. Study on the mechanism of action of artemether against schistosomes: The identification of cysteine adducts of both carbon-centred free radicals derived from artemether. Bioorg. Med. Chem. Lett. 2003, 13, 1645–1647.
- Eckstein-Ludwig, U.; Webb, R.J.; Van Goethem, I.D.A.; East, J.M.; Lee, A.G.; Kimura, M.; O’Neill, P.M.; Bray, P.G.; Ward, S.A.; Krishna, S. Artemisinins target the SERCA of Plasmodium falciparum. Nature 2003, 424, 957–961.
- Wang, J.; Xu, C.; Lun, Z.R.; Meshnick, S.R. Unpacking ‘Artemisinin Resistance’. Trends Pharmacol. Sci. 2017, 38, 506–511.
- Jena, N.R. DNA damage by reactive species: Mechanisms, mutation and repair. J. Biosci. 2012, 37, 503–517.
- Cadet, J.; Wagner, J.R. DNA base damage by reactive oxygen species, oxidizing agents, and UV radiation. Cold Spring Harb. Perspect. Biol. 2013, 5, a012559.
- Gopalakrishnan, A.M.; Kumar, N. Antimalarial action of artesunate involves DNA damage mediated by reactive oxygen species. Antimicrob. Agents Chemother. 2015, 59, 317–325.
- Uhlemann, A.C.; Wittlin, S.; Matile, H.; Bustamante, L.Y.; Krishna, S. Mechanism of antimalarial action of the synthetic trioxolane RBX11160 (OZ277). Antimicrob. Agents Chemother. 2007, 51, 667–672.
- Postma, N.S.; Mommers, E.C.; Eling, W.M.C.; Zuidema, J. Oxidative stress in malaria; implications for prevention and therapy. Pharm. World Sci. 1996, 18, 121–129.
- Rahbari, M.; Rahlfs, S.; Jortzik, E.; Bogeski, I.; Becker, K. H2O2 dynamics in the malaria parasite Plasmodium falciparum. PLoS ONE 2017, 12, e0174837.
- Percário, S.; Moreira, D.R.; Gomes, B.A.Q.; Ferreira, M.E.S.; Gonçalves, A.C.M.; Laurindo, P.S.O.C.; Vilhena, T.C.; Dolabela, M.F.; Green, M.D. Oxidative stress in Malaria. Int. J. Mol. Sci. 2012, 13, 16346–16372.
- Tiwari, S.; Sharma, N.; Sharma, G.P.; Mishra, N. Redox interactome in malaria parasite Plasmodium falciparum. Parasitol. Res. 2021, 120, 423–434.
- Loria, P.; Miller, S.; Foley, M.; Tilley, L. Inhibition of the peroxidative degradation of haem as the basis of action of chloroquine and other quinoline antimalarials. Biochem. J. 1999, 339, 363–370.
- Zhang, J.; Krugliak, M.; Ginsburg, H. The fate of ferriprotorphyrin IX in malaria infected erythrocytes in conjunction with the mode of action of antimalarial drugs. Mol. Biochem. Parasitol. 1999, 99, 129–141.
- Egan, T.J.; Combrinck, J.M.; Egan, J.; Hearne, G.R.; Marques, H.M.; Ntenteni, S.; Sewell, B.T.; Smith, P.J.; Taylor, D.; Van Schalkwyk, D.A.; et al. Fate of haem iron in the malaria parasite Plasmodium falciparum. Biochem. J. 2002, 365, 343–347.
- Rahlfs, S.; Schirmer, R.H.; Becker, K. The thioredoxin system of Plasmodium falciparum and other parasites. Cell. Mol. Life Sci. 2002, 59, 1024–1041.
- Desakorn, V.; Dondorp, A.M.; Silamut, K.; Pongtavornpinyo, W.; Sahassananda, D.; Chotivanich, K.; Pitisuttithum, P.; Smithyman, A.M.; Day, N.P.J.; White, N.J. Stage-dependent production and release of histidine-rich protein 2 by Plasmodium falciparum. Trans. R. Soc. Trop. Med. Hyg. 2005, 99, 517–524.
- Wrenger, C.; Eschbach, M.L.; Müller, I.B.; Warnecke, D.; Walter, R.D. Analysis of the vitamin B6 biosynthesis pathway in the human malaria parasite Plasmodium falciparum. J. Biol. Chem. 2005, 280, 5242–5248.
- Nickel, C.; Rahlfs, S.; Deponte, M.; Koncarevic, S.; Becker, K. Thioredoxin networks in the malarial parasite Plasmodium falciparum. Antioxid. Redox Signal. 2006, 8, 1227–1239.
- Egwu, C.O.; Tsamesidis, I.; Pério, P.; Augereau, J.-M.; Benoit-Vical, F.; Reybier, K. Superoxide: A major role in the mechanism of action of essential antimalarial drugs. Free Radic. Biol. Med. 2021, 167, 271–275.
- Tsamesidis, I.; Egwu, C.O.; Pério, P.; Augereau, J.M.; Benoit-Vical, F.; Reybier, K. An LC–MS assay to measure superoxide radicals and hydrogen peroxide in the blood system. Metabolites 2020, 10, 175.
- Sullivan, D.J.; Matile, H.; Ridley, R.G.; Goldberg, D.E. A common mechanism for blockade of heme polymerization by antimalarial quinolines. J. Biol. Chem. 1998, 273, 31103–31107.
- Sugioka, Y.; Suzuki, M.; Sugioka, K.; Nakano, M. A ferriprotoporphyrin IX-chloroquine complex promotes membrane phospholipid peroxidation A possible mechanism for antimalarial action. FEBS Lett. 1987, 223, 251–254.
- Haynes, R.K.; Cheu, K.W.; Chan, H.W.; Wong, H.N.; Li, K.Y.; Tang, M.M.K.; Chen, M.J.; Guo, Z.F.; Guo, Z.H.; Sinniah, K.; et al. Interactions between Artemisinins and other Antimalarial Drugs in Relation to the Cofactor Model-A Unifying Proposal for Drug Action. ChemMedChem 2012, 7, 2204–2226.
- De Almeida Ribeiro, M.C.; Augusto, O.; Da Costa Ferreira, A.M. Inhibitory effect of chloroquine on the peroxidase activity of ferriprotoporphyrin IX. J. Chem. Soc. Dalt. Trans. 1995, 3759–3766.
- Birth, D.; Kao, W.C.; Hunte, C. Structural analysis of atovaquone-inhibited cytochrome bc 1 complex reveals the molecular basis of antimalarial drug action. Nat. Commun. 2014, 5, 1–11.
- Fry, M.; Pudney, M. Site of action of the antimalarial hydroxynaphthoquinone, 2--3- hydroxy-1,4-naphthoquinone (566C80). Biochem. Pharmacol. 1992, 43, 1545–1553.
- Srivastava, I.K.; Rottenberg, H.; Vaidya, A.B. Atovaquone, a broad spectrum antiparasitic drug, collapses mitochondrial membrane potential in a malarial parasite. J. Biol. Chem. 1997, 272, 3961–3966.
- Barton, V.; Fisher, N.; Biagini, G.A.; Ward, S.A.; O’Neill, P.M. Inhibiting Plasmodium cytochrome bc1: A complex issue. Curr. Opin. Chem. Biol. 2010, 14, 440–446.
- Fiorillo, M.; Lamb, R.; Tanowitz, H.B.; Mutti, L.; Krstic-Demonacos, M.; Cappello, A.R.; Martinez-Outschoorn, U.E.; Sotgia, F.; Lisanti, M.P. Repurposing atovaquone: Targeting mitochondrial complex III and OXPHOS to eradicate cancer stem cells. Oncotarget 2016, 7, 34084–34099.
- Posner, G.H.; Oh, C.H. A Regiospecifically Oxygen-18 Labeled 1,2,4-Trioxane: A Simple Chemical Model System To Probe the Mechanism(s) for the Antimalarial Activity of Artemisinin (Qinghaosu). J. Am. Chem. Soc. 1992, 114, 8328–8329.
- Mercer, A.E.; Copple, I.M.; Maggs, J.L.; O’Neill, P.M.; Park, B.K. The role of heme and the mitochondrion in the chemical and molecular mechanisms of mammalian cell death induced by the artemisinin antimalarials. J. Biol. Chem. 2011, 286, 987–996.
- Posner, G.H.; Oh, C.H.; Wang, D.; Gerena, L.; Milhous, W.K.; Meshnick, S.R.; Asawamahasadka, W. Mechanism-Based Design, Synthesis, and in Vitro Antimalarial Testing of New 4-Methylated Trioxanes Structurally Related to Artemisinin: The Importance of a Carbon-Centered Radical for Antimalarial Activity. J. Med. Chem. 1994, 37, 1256–1258.
- Robert, A.; Benoit-Vical, F.; Claparols, C.; Meunier, B. The antimalarial drug artemisinin alkylates heme in infected mice. Proc. Natl. Acad. Sci. USA 2005, 102, 13676–13680.
- Wang, J.; Huang, L.; Li, J.; Fan, Q.; Long, Y.; Li, Y.; Zhou, B. Artemisinin directly targets malarial mitochondria through its specific mitochondrial activation. PLoS ONE 2010, 5, e9582.
- Allen, R.J.W.; Kirk, K. The Membrane Potential of the Intraerythrocytic Malaria Parasite Plasmodium falciparum. J. Biol. Chem. 2004, 279, 11264–11272.
- Biagini, G.A.; Viriyavejakul, P.; O’Neill, P.M.; Bray, P.G.; Ward, S.A. Functional characterization and target validation of alternative complex I of Plasmodium falciparum mitochondria. Antimicrob. Agents Chemother. 2006, 50, 1841–1851.
- Antoine, T.; Fisher, N.; Amewu, R.; O’Neill, P.M.; Ward, S.A.; Biagini, G.A. Rapid kill of malaria parasites by artemisinin and semi-synthetic endoperoxides involves ROS-dependent depolarization of the membrane potential. J. Antimicrob. Chemother. 2014, 69, 1005–1016.
- Egwu, C.O.; Pério, P.; Augereau, J.-M.; Tsamesidis, I.; Benoit-Vical, F.; Reybier, K. Resistance to artemisinin in falciparum malaria parasites: A redox-mediated phenomenon. Free Radic. Biol. Med. 2021, S0891-5849, 00476–00477.
- WHO. Artemisinin Resistance and Artemisinin-Based Combination Therapy Efficacy (Status Report—August 2018). Available online: https://apps.who.int/iris/bitstream/handle/10665/274362/WHO-CDS-GMP-2018.18-eng.pdf?ua=1 (accessed on 26 December 2020).
- Van Vugt, M.; Brockman, A.; Gemperli, B.; Luxemburger, C.; Gathmann, I.; Royce, C.; Slight, T.; Looareesuwan, S.; White, N.J.; Nosten, F. Randomized comparison of artemether-benflumetol and artesunate-mefloquine in treatment of multidrug-resistant falciparum malaria. Antimicrob. Agents Chemother. 1998, 42, 135–139.
- Rudrapal, M.; Chetia, D. Endoperoxide antimalarials: Development, structural diversity and pharmacodynamic aspects with reference to 1,2,4-trioxane-based structural scaffold. Drug Des. Devel. Ther. 2016, 10, 3575–3590.
- Benoit-Vical, F.; Lelièvre, J.; Berry, A.; Deymier, C.; Dechy-Cabaret, O.; Cazelles, J.; Loup, C.; Robert, A.; Magnaval, J.F.; Meunier, B. Trioxaquines are new antimalarial agents active on all erythrocytic forms, including gametocytes. Antimicrob. Agents Chemother. 2007, 51, 1463–1472.
- Toler, S. The plasmodial apicoplast was retained under evolutionary selective pressure to assuage blood stage oxidative stress. Med. Hypotheses 2005, 65, 683–690.
- Storm, J. Lipoic Acid Metabolism of Plasmodium—A Suitable Drug Target. Curr. Pharm. Des. 2012, 18, 3480.
- Birnbaum, J.; Scharf, S.; Schmidt, S.; Jonscher, E.; Maria Hoeijmakers, W.A.; Flemming, S.; Toenhake, C.G.; Schmitt, M.; Sabitzki, R.; Bergmann, B.; et al. A Kelch13-defined endocytosis pathway mediates artemisinin resistance in malaria parasites. Science 2020, 367, 51–59.
- Zhang, S.; Gerhard, G.S. Heme activates artemisinin more efficiently than hemin, inorganic iron, or hemoglobin. Bioorg. Med. Chem. 2008, 16, 7853–7861.
- Xie, S.C.; Dogovski, C.; Hanssen, E.; Chiu, F.; Yang, T.; Crespo, M.P.; Stafford, C.; Batinovic, S.; Teguh, S.; Charman, S.; et al. Haemoglobin degradation underpins the sensitivity of early ring stage Plasmodium falciparum to artemisinins. J. Cell Sci. 2016, 129, 406–416.
- Koncarevic, S.; Rohrbach, P.; Deponte, M.; Krohne, G.; Prieto, J.H.; Yates, J.; Rahlfs, S.; Becker, K. The malarial parasite Plasmodium falciparum imports the human protein peroxiredoxin 2 for peroxide detoxification. Proc. Natl. Acad. Sci. USA 2009, 106, 13323–13328.
- Fairfield, A.S.; Meshnick, S.R.; Eaton, J.W. Malaria parasites adopt host cell superoxide dismutase. Science 1983, 221, 764–766.
- Clarebout, G.; Slomianny, C.; Delcourt, P.; Leu, B.; Masset, A.; Camus, D.; Dive, D. Status of Plasmodium falciparum towards catalase. Br. J. Haematol. 1998, 103, 52–59.
- Varadharajan, S.; Sagar, B.K.C.; Rangarajan, P.N.; Padmanaban, G. Localization of ferrochelatase in Plasmodium falciparum. Biochem. J. 2004, 384, 429–436.
- Bonday, Z.Q.; Dhanasekaran, S.; Rangarajan, P.N.; Padmanaban, G. Import of host δ-aminolevulinate dehydratase into the malarial parasite: Identification of a new drug target. Nat. Med. 2000, 6, 898–903.
- Nogueira, F.; Diez, A.; Radfar, A.; Pérez-Benavente, S.; Rosario, V.E.; Puyet, A.; Bautista, J.M. Early transcriptional response to chloroquine of the Plasmodium falciparum antioxidant defence in sensitive and resistant clones. Acta Trop. 2010, 114, 109–115.
- Rocamora, F.; Zhu, L.; Liong, K.Y.; Dondorp, A.; Miotto, O.; Mok, S.; Bozdech, Z. Oxidative stress and protein damage responses mediate artemisinin resistance in malaria parasites. PLoS Pathog. 2018, 14, e1006930.
- Mok, S.; Ashley, E.A.; Ferreira, P.E.; Zhu, L.; Lin, Z.; Yeo, T.; Chotivanich, K.; Imwong, M.; Pukrittayakamee, S.; Dhorda, M.; et al. Population transcriptomics of human malaria parasites reveals the mechanism of artemisinin resistance. Science 2015, 347, 431–435.
- Mbengue, A.; Bhattacharjee, S.; Pandharkar, T.; Liu, H.; Estiu, G.; Stahelin, R.V.; Rizk, S.S.; Njimoh, D.L.; Ryan, Y.; Chotivanich, K.; et al. A molecular mechanism of artemisinin resistance in Plasmodium falciparum malaria. Nature 2015, 520, 683–687.
- Tawk, L.; Chicanne, G.; Dubremetz, J.F.; Richard, V.; Payrastre, B.; Vial, H.J.; Roy, C.; Wengelnik, K. Phosphatidylinositol 3-phosphate, an essential lipid in Plasmodium, localizes to the food vacuole membrane and the apicoplast. Eukaryot. Cell 2010, 9, 1519–1530.
- Ariey, F.; Witkowski, B.; Amaratunga, C.; Beghain, J.; Langlois, A.C.; Khim, N.; Kim, S.; Duru, V.; Bouchier, C.; Ma, L.; et al. A molecular marker of artemisinin-resistant Plasmodium falciparum malaria. Nature 2014, 505, 50–55.
- Isozumi, R.; Uemura, H.; Kimata, I.; Ichinose, Y.; Logedi, J.; Omar, A.H.; Kaneko, A. Novel mutations in k13 propeller gene of artemisinin-resistant Plasmodium falciparum. Emerg. Infect. Dis. 2015, 21, 490–492.
- Balikagala, B.; Mita, T.; Ikeda, M.; Sakurai, M.; Yatsushiro, S.; Takahashi, N.; Tachibana, S.I.; Auma, M.; Ntege, E.H.; Ito, D.; et al. Absence of in vivo selection for K13 mutations after artemether-lumefantrine treatment in Uganda. Malar. J. 2017, 16, 23.
- Lee, A.H.; Fidock, D.A. Evidence of a mild mutator phenotype in cambodian Plasmodium falciparum malaria parasites. PLoS ONE 2016, 11, e0154166.