Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Subjects:
Biochemical Research Methods
|
Agriculture, Dairy & Animal Science
|
Biotechnology & Applied Microbiology
The Pseudomonas genus is ubiquitous and comprises species which are well known phytopathogens, such as P. syringae, or opportunistic human pathogens, such as P. aeruginosa, but also host members associated with water, soil and plant surfaces. Pseudomonas spp. are well adapted to growing in the rhizosphere and are well suited for biocontrol and growth promotion. Pseudomonas lipopeptides (Ps-LPs) play crucial roles in bacterial physiology, host–microbe interactions and plant disease control.
- Pseudomonas fluorescens
- lipopeptides
- antimicrobial
- plant-microbe interactions
- direct antagonism
- induced systemic resistance
- secondary metabolites
1. Introduction
The Pseudomonas genus is ubiquitous and comprises species which are well known phytopathogens, such as P. syringae, or opportunistic human pathogens, such as P. aeruginosa, but also host members associated with water, soil and plant surfaces [1]. Pseudomonas spp. are well adapted to growing in the rhizosphere and are well suited for biocontrol and growth promotion [2]. Thus, the use of fluorescent Pseudomonas spp. as potential biopesticides has gained attention over the last decade. These bacteria are of particular interest because of their enormous metabolic versatility and wide adaptation across environmental gradients [3].
Based on phylogenomic and Multi Locus Sequence Analyses (MLSA), the Pseudomonas genus has been delineated into 453 species (https://lpsn.dsmz.de/genus/pseudomonas; accessed on 18 December 2021) which are distributed across three lineages (P. fluorescens, P. aeruginosa and P. pertucinogena), several groups (G) and subgroups (SG) [4][5][6][7][8]. Most biocontrol strains have been described within the P. fluorescens group comprising among others, the P. fluorescens SG, P. koreensis SG, P. chlororaphis SG, P. jessenii SG, P. mandelii SG and P. corrugata SG. Additionally, several biocontrol strains are positioned within the P. putida and P. syringae groups. These disease-suppressing pseudomonads were isolated from several sources ranging from the healthy plant rhizosphere [9][10][11], plant rhizosphere [12][13][14][15], phyllosphere [16][17], bulk soil [15] and suppressive soils [10][18]. The commonality among well-studied biocontrol strains is their capacity for secondary metabolite production including siderophores, lipopeptides (LPs), hydrogen cyanide, bacteriocins and certain antibiotics such as phenazines, 2,4-diacetylphloroglucinol (DAPG), pyrrolnitrin and pyoluteorin [3][19].
Examples of commercially available Pseudomonas-based bioprotectants include fungicides such as Cedomon and Cerall (P. chlororaphis MA342) both targeting seed-borne pathogens of cereals, Spot-Less (P. aureofaciens strain Tx-1) for management of fungal diseases on lawns and grasses, and Howler (P. chlororaphis AFS009) useful in the management of Rhizoctonia, Pythium, Fusarium, Phytophthora, Colletotrichum spp. in fruits, vegetables and ornamentals [19]. A detailed list of commercial bioprotectants based on Pseudomonas in Europe and USA, including their usage, and target crops/applications/pathogens have been enumerated in a recent review [19].
Lipopeptides are bacterial metabolites consisting of a peptide part attached to a fatty acid tail [1]. Most beneficial LPs are cyclized although linear LPs have also been described [20][21]. LPs have drawn remarkable interest because of their broad-spectrum antimicrobial and ecological functions. These multiple functions include biofilm formation and colonization of surfaces, quorum sensing, cell motility, soil remediation, anti-oomycete, antiviral, antifungal, antibacterial, herbicidal, insecticidal, antiprotozoal and anticancer properties [3][22][23][24][25][26][27].
2. Genome Comparison of Selected Lipopeptide-Producing Pseudomonas spp.
A previous study provided the phylogenomic analysis of the Pseudomonas genus based on the genomes of the type strains of 163 described species and compared these type strain genomes to those of 1223 Pseudomonas genomes in public databases [7]. Results showed that 400 of those 1223 genomes were distinct from any other type strain suggesting that the Pseudomonas genomic diversity had been grossly underrepresented by the type strains. Furthermore, a detailed comparative genome analysis of ten strains within the Pseudomonas fluorescens group highlighted the enormous diverseness of this group and the capacity of the variable genome to adapt individual strains to their distinct lifestyles and functional capacities [3]. Here, using the P. fluorescens Pf0-1 as a reference genome, we compared the genome of 32 lipopeptide-producing Pseudomonas strains affiliated with the P. koreensis, P. fluorescens, P. mandelii, P. corrugata, P. asplenii, P. chlororaphis, P. protegens, subgroups including the P. putida and P. syringae groups. By comparing the protein coding sequences (CDS) of reference to query genomes, a Blast Atlas was generated which showed the close relatedness of other members of the P. koreensis group (P. fluorescens MS80, P. granadensis LMG 27,940 and P. kribbensis 46-2) to the reference genome P. fluorescens Pf0-1 (Figure 3).
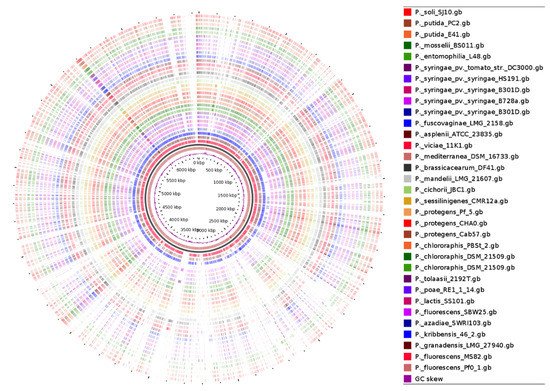
Figure 3. Comparative Genome Blast Atlas of 35 Lipopeptide-Producing Pseudomonas Strains. The BLAST Atlas analysis displays regions of the uploaded query files (34 genomes) where there are BLAST hits to the reference genome P. fluorescens Pf0-1). The GView Server was used [28].
3. Chemical Diversity of Beneficial Pseudomonas LPs
Most beneficial LPs have been predominantly characterized from strains affiliated with the P. fluorescens and P. putida group. The chemical diversity of Pseudomonas LPs has been detailed in two recent reviews [1][21]. Table 1 shows the diversity of beneficial LPs and presents the discovery of similar LPs from diverse strains, countries, niches and environments. Not all LPs listed have been functionally characterized, however, the disease suppressive capacity of their producing strain(s) has been established on specific plant hosts thus indicating non-virulence. Clearly, the P. koreensis subgroup presents the highest diversity of LP families and individual members, including variants. This SG is characterized by at least six amphisin group members alongside the novel rhizoamide, the bananamide group comprising six variants and the cocoyamide/gacamide group. Moderate LP diversity is showcased by the P. fluorescens SG while the P. protegens SG comprises various orfamide variants A-H and the poaeamide LPs. Lastly, the P. putida group contains four described LP types: entolysin, putisolvin, xantholysin, WLIP and a novel 17AA LP named N8. Figure 4 shows the chemical structures of representative biocontrol LPs that have been characterized.

Figure 4. Chemical structures of selected biologically active Pseudomonas Cyclic Lipopeptides. Bananamide D (Bananamide Group); WLIP (Viscosin Group); Thanamycin (Syringomycin Group); Lokisin (Amphisin Group); Cocoyamide; Putisolvin I; Entolysin A and Xantholysin A. Whenever the absolute configuration of the lipopeptides was reported in the literature, it is indicated by standard stereodescriptors. In case of WLIP, the 3D-structure was secured by x-ray [29] and can be viewed as entry CCDC 919,229 at The Cambridge Crystallographic Data Centre via www.ccdc.cam.ac.uk (accessed on 19 December 2021).
Table 1. Taxonomy of LP-producing Biocontrol Pseudomonads, their corresponding Molecules and Origin.
| Taxonomy | Biocontrol Strains | Host/Origin | Country | LP Family | LP | Reference |
|---|---|---|---|---|---|---|
| P. fluorescens SG | SS101 | Wheat rhizosphere | Netherlands | Viscosin | Massetolide | [30] |
| SBW25 | Sugarbeet phyllosphere | UK | Viscosin | [31] | ||
| DR54 | Sugarbeet rhizosphere | Denmark | Viscosinamide | [32][33] | ||
| A2W4.9, U2W1.5 | White cocoyam rhizosphere | Nigeria | Viscosinamide | [34] | ||
| BRG100 | Green foxtail rhizosphere | Canada | Pseudophomin | [35] | ||
| RE*1-1-14 | Internal part of soybean roots | Germany | Poaemide | [36][37] | ||
| NCPPB1311 | Cultivated mushrooms | UK | WLIP | [38] | ||
| P. koreensis SG | DSS73 | Sugarbeet rhizosphere | Denmark | Amphisin | Amphisin | [14][39] |
| HKI0770 | Forest soil | Forest soil | Anikasin | [40][41] | ||
| CTS17 | Sugarbeet rhizosphere | Denmark | Hodersin | [14][42] | ||
| DSS41 | Sugarbeet rhizosphere | Denmark | Lokisin | [43] | ||
| 2.74 | Tomato hydroponics | Sweden | Lokisin | [44] | ||
| S150 | Tobacco rhizosphere | China | Lokisin | [45] | ||
| COR10 | Red cocoyam rhizosphere | Cameroon | Lokisin | [10] | ||
| UCMA 17988 | Raw bulk tank milk | France | Milkisin | [46] | ||
| COW8 | White cocoyam rhizosphere | Cameroon | Rhizoamide (N2—11:7) † | [11] | ||
| 96.578 | Sugarbeet rhizosphere | Denmark | Tensin | [33][47] | ||
| BW11P2 | Banana rhizoplane | Sri Lanka | Bananamide | Bananamide I, II, III | [12][48] | |
| COW3, COW65 | White cocoyam rhizosphere | Cameroon | Bananamide D, E, F, G | [10][49] | ||
| COW5 | White cocoyam rhizosphere | Cameroon | Cocoyamide | Cocoyamide A | [10] | |
| Pf0-1 | Loam soil | USA | Gacamide A | [50][51] | ||
| P. protegens SG | CHA0 | Tobacco roots | Switzerland | Orfamide | Orfamide | [52][53] |
| Pf-5 | Cotton rhizosphere | USA | Orfamide | [54][55] | ||
| CMR5c | Red cocoyam rhizosphere | Cameroon | Orfamide | [53] | ||
| CMR12a | Red cocoyam rhizosphere | Cameroon | Orfamide, Sessilin | [56] | ||
| P. chlororaphis SG | COR52 | Red cocoyam rhizosphere | Cameroon | Viscosin | Pseudodesmin | [34] |
| P. mandelii SG | In5 | Suppressive potato soil | Greenland | Syringomycin | Nunamycin | [18] |
| In5 | Suppressive potato soil | Greenland | Syringopeptin | Nunapeptin | [18] | |
| P. corrugata SG | SH-C52 | Sugarbeet rhizosphere | Netherlands | Syringomycin | Thanamycin | [57] |
| DF41 | Canola root | Canada | Thanamycin -var1 | [58][59] | ||
| 11K1 | Bean rhizosphere | China | Brasmycin | [60] | ||
| SH-C52 | Sugarbeet rhizosphere | Netherlands | Syringopeptin | Thanapeptin | [57] | |
| DF41 | Canola root | Canada | Sclerosin | [59] | ||
| 11K1 | Bean rhizosphere | China | Braspeptin | [60] | ||
| P. putida G | BW11M1 | Banana rhizoplane | Sri Lanka | Xantholysin | Xantholysin | [12][61] |
| COR51 | Red cocoyam rhizosphere | Cameroon | Xantholysin | [10] | ||
| BS011 | Rice rhizosphere | China | Xantholysin | [62] | ||
| 267 | Black pepper | Vietnam | Putisolvin | Putisolvin I, II | [63] | |
| COR55 | Red cocoyam rhizosphere | Cameroon | Putisolvin III, IV, V | [10][11] | ||
| L48 | Fly | Guadeloupe | Entolysin | Entolysin A, B | [64] | |
| COR5 | Red cocoyam rhizosphere | Cameroon | Entolysin B | [10] | ||
| RW10S2 | Rice rhizosphere | Sri Lanka | Viscosin | WLIP | [65] | |
| COW10 | White cocoyam rhizosphere | Cameroon | WLIP | [10] | ||
| NSE1 | White cocoyam rhizosphere | Nigeria | WLIP | [66] | ||
| COR35 | Red cocoyam rhizosphere | Cameroon | Unclassified | N8 (17:8) † | [11] | |
| P. asplenii SG | COR33 | Red cocoyam rhizosphere | Cameroon | Unclassified | N5 (13:8) † | [11] |
| COR18 | Red cocoyam rhizosphere | Cameroon | N5 (13:8), N7 †, Mycin LP † | [11] | ||
| Novel U2 SG | COR58 | Red cocoyam rhizosphere | Cameroon | Unclassified | N4 (12:10) † | [10][11] |
† novel LPs.
This entry is adapted from the peer-reviewed paper 10.3390/molecules27020372
References
- Götze, S.; Stallforth, P. Structure, properties, and biological functions of nonribosomal lipopeptides from pseudomonads. Nat. Prod. Rep. 2020, 37, 29–54.
- Weller, D.M. Pseudomonas Biocontrol Agents of Soilborne Pathogens: Looking Back Over 30 Years. Phytopathology 2007, 97, 250–256.
- Loper, J.E.; Hassan, K.A.; Mavrodi, D.V.; Davis, E.W., II; Lim, C.K.; Shaffer, B.T.; Elbourne, L.D.H.; Stockwell, V.O.; Hartney, S.L.; Breakwell, K.; et al. Comparative Genomics of Plant-Associated Pseudomonas spp.: Insights into Diversity and Inheritance of Traits Involved in Multitrophic Interactions. PLoS Genet. 2012, 8, e1002784.
- Lalucat, J.; Mulet, M.; Gomila, M.; García-Valdés, E. Genomics in Bacterial Taxonomy: Impact on the Genus Pseudomonas. Genes 2020, 11, 139.
- Gomila, M.; Peña, A.; Mulet, M.; Lalucat, J.; García-Valdés, E. Phylogenomics and systematics in Pseudomonas. Front. Microbiol. 2015, 6, 214.
- Girard, L.; Lood, C.; Höfte, M.; Vandamme, P.; Rokni-Zadeh, H.; van Noort, V.; Lavigne, R.; De Mot, R. The Ever-Expanding Pseudomonas Genus: Description of 43 New Species and Partition of the Pseudomonas putida Group. Microorganisms 2021, 9, 1766.
- Hesse, C.; Schulz, F.; Bull, C.T.; Shaffer, B.T.; Yan, Q.; Shapiro, N.; Hassan, K.A.; Varghese, N.; Elbourne, L.D.H.; Paulsen, I.T.; et al. Genome-based evolutionary history of Pseudomonas spp. Environ. Microbiol. 2018, 20, 2142–2159.
- Behzadi, P.; Baráth, Z.; Gajdács, M. It’s Not Easy Being Green: A Narrative Review on the Microbiology, Virulence and Therapeutic Prospects of Multidrug-Resistant Pseudomonas aeruginosa. Antibiotics 2021, 10, 42.
- Perneel, M.; Heyrman, J.; Adiobo, A.; De Maeyer, K.; Raaijmakers, J.M.; De Vos, P.; Höfte, M. Characterization of CMR5c and CMR12a, novel fluorescent Pseudomonas strains from the cocoyam rhizosphere with biocontrol activity. J. Appl. Microbiol. 2007, 103, 1007–1020.
- Oni, F.E.; Geudens, N.; Omoboye, O.O.; Bertier, L.; Hua, H.G.K.; Adiobo, A.; Sinnaeve, D.; Martins, J.C.; Höfte, M. Fluorescent Pseudomonas and cyclic lipopeptide diversity in the rhizosphere of cocoyam (Xanthosoma sagittifolium). Environ. Microbiol. 2019, 21, 1019–1034.
- Oni, F.E.; Geudens, N.; Onyeka, J.T.; Olorunleke, O.F.; Salami, A.E.; Omoboye, O.O.; Arias, A.A.; Adiobo, A.; De Neve, S.; Ongena, M.; et al. Cyclic lipopeptide-producing Pseudomonas koreensis group strains dominate the cocoyam rhizosphere of a Pythium root rot suppressive soil contrasting with P. putida prominence in conducive soils. Environ. Microbiol. 2020, 22, 5137–5155.
- Vlassak, K.; Van Holm, L.; Duchateau, L.; Vanderleyden, J.; De Mot, R. Isolation and characterization of fluorescent Pseudomonas associated with the roots of rice and banana grown in Sri Lanka. Plant Soil 1992, 145, 51–63.
- Tran, H.; Kruijt, M.; Raaijmakers, J.M. Diversity and activity of biosurfactant-producing Pseudomonas in the rhizosphere of black pepper in Vietnam. J. Appl. Microbiol. 2008, 104, 839–851.
- Nielsen, T.H.; Sørensen, D.; Tobiasen, C.; Andersen, J.B.; Christophersen, C.; Givskov, M.; Sørensen, J. Antibiotic and Biosurfactant Properties of Cyclic Lipopeptides Produced by Fluorescent Pseudomonas spp. from the Sugar Beet Rhizosphere. Appl. Environ. Microbiol. 2002, 68, 3416–3423.
- Lopes, L.D.; Davis, E.W.; Pereira e Silva, M.d.C.; Weisberg, A.J.; Bresciani, L.; Chang, J.H.; Loper, J.E.; Andreote, F.D. Tropical soils are a reservoir for fluorescent Pseudomonas spp. biodiversity. Environ. Microbiol. 2018, 20, 62–74.
- Berry, C.; Fernando, W.G.D.; Loewen, P.C.; de Kievit, T.R. Lipopeptides are essential for Pseudomonas sp. DF41 biocontrol of Sclerotinia sclerotiorum. Biol. Control. 2010, 55, 211–218.
- Müller, T.; Behrendt, U.; Ruppel, S.; Von Der Waydbrink, G.; Müller, M.E.H. Fluorescent Pseudomonads in the Phyllosphere of Wheat: Potential Antagonists Against Fungal Phytopathogens. Curr. Microbiol. 2015, 72, 383–389.
- Michelsen, C.F.; Watrous, J.; Glaring, M.A.; Kersten, R.; Koyama, N.; Dorrestein, P.C.; Stougaard, P. Nonribosomal Peptides, Key Biocontrol Components for Pseudomonas fluorescens In5, Isolated from a Greenlandic Suppressive Soil. mBio 2015, 6, e00079.
- Höfte, M. The Use of Pseudomonas spp. as Bacterial Biocontrol Agents to Control Plant Disease. In Microbial Bioprotectants for Plant Disease Management; Burleigh Dodds Science Publishing: Cambridge, UK, 2021; p. 75.
- Olorunleke, F.E.; Kieu, N.P.; Höfte, M. Recent Advances in Pseudomonas Biocontrol. In Bacterial-Plant Interactions: Advance Research and Future Trends; Caister Academic Press: Cambridge, UK, 2015; Volume 1, pp. 167–198.
- Geudens, N.; Martins, J.C. Cyclic Lipodepsipeptides From Pseudomonas spp.—Biological Swiss-Army Knives. Front. Microbiol. 2018, 9, 1–18.
- Raaijmakers, J.M.; De Bruijn, I.; Nybroe, O.; Ongena, M. Natural functions of lipopeptides from Bacillus and Pseudomonas: More than surfactants and antibiotics. FEMS Microbiol. Rev. 2010, 34, 1037–1062.
- Calderón, C.E.; Pérez-García, A.; de Vicente, A.; Cazorla, F.M. The dar Genes of Pseudomonas chlororaphis PCL1606 Are Crucial for Biocontrol Activity via Production of the Antifungal Compound 2-Hexyl, 5-Propyl Resorcinol. Mol. Plant Microbe Interact. 2013, 26, 554–565.
- Flury, P.; Aellen, N.; Ruffner, B.; Péchy-Tarr, M.; Fataar, S.; Metla, Z.; Ferreras, A.D.; Bloemberg, G.; Frey, J.; Goesmann, A.; et al. Insect pathogenicity in plant-beneficial pseudomonads: Phylogenetic distribution and comparative genomics. ISME J. 2016, 10, 2527–2542.
- Flury, P.; Vesga, P.; Péchy-Tarr, M.; Aellen, N.; Dennert, F.; Hofer, N.; Kupferschmied, K.P.; Kupferschmied, P.; Metla, Z.; Ma, Z.; et al. Antimicrobial and Insecticidal: Cyclic Lipopeptides and Hydrogen Cyanide Produced by Plant-Beneficial Pseudomonas Strains CHA0, CMR12a, and PCL1391 Contribute to Insect Killing. Front. Microbiol. 2017, 8, 100.
- Garrido-Sanz, D.; Meier-Kolthoff, J.P.; Göker, M.; Martín, M.; Rivilla, R.; Redondo-Nieto, M. Genomic and Genetic Diversity within the Pseudomonas fluorescens Complex. PLoS ONE 2016, 11, e0150183.
- Garrido-Sanz, D.; Arrebola, E.; Martínez-Granero, F.; García-Méndez, S.; Muriel, C.; Blanco-Romero, E.; Martín, M.; Rivilla, R.; Redondo-Nieto, M. Classification of Isolates from the Pseudomonas fluorescens Complex into Phylogenomic Groups Based in Group-Specific Markers. Front. Microbiol. 2017, 8, 413.
- Petkau, A.; Stuart-Edwards, M.; Stothard, P.; Van Domselaar, G. Interactive microbial genome visualization with GView. Bioinformatics 2010, 26, 3125–3126.
- Mehnaz, S.; Saleem, R.S.Z.; Yameen, B.; Pianet, I.; Schnakenburg, G.; Pietraszkiewicz, H.; Valeriote, F.; Josten, M.; Sahl, H.-G.; Franzblau, S.G.; et al. Lahorenoic Acids A–C, ortho-Dialkyl-Substituted Aromatic Acids from the Biocontrol Strain Pseudomonas aurantiaca PB-St2. J. Nat. Prod. 2013, 76, 135–141.
- de Souza, J.T.; de Boer, M.; de Waard, P.; van Beek, T.A.; Raaijmakers, J.M. Biochemical, Genetic, and Zoosporicidal Properties of Cyclic Lipopeptide Surfactants Produced by Pseudomonas fluorescens. Appl. Environ. Microbiol. 2003, 69, 7161–7172.
- de Bruijn, I.; de Kock, M.J.D.; Yang, M.; de Waard, P.; van Beek, T.A.; Raaijmakers, J.M. Genome-based discovery, structure prediction and functional analysis of cyclic lipopeptide antibiotics in Pseudomonas species. Mol. Microbiol. 2007, 63, 417–428.
- Thrane, C.; Olsson, S.; Nielsen, T.H.; Sørensen, J. Vital fluorescent stains for detection of stress in Pythium ultimum and Rhizoctonia solani challenged with viscosinamide from Pseudomonas fluorescens DR54. FEMS Microbiol. Ecol. 1999, 30, 11–23.
- Nielsen, M.N.; Sørensen, J.; Fels, J.; Pedersen, H.C. Secondary Metabolite- and Endochitinase-Dependent Antagonism toward Plant-Pathogenic Microfungi of Pseudomonas fluorescens Isolates from Sugar Beet Rhizosphere. Appl. Environ. Microbiol. 1998, 64, 3563–3569.
- Oni, F.E.; Geudens, N.; Adiobo, A.; Omoboye, O.O.; Enow, E.A.; Onyeka, J.T.; Salami, A.E.; De De Mot, R.; Martins, J.C.; Höfte, M. Biosynthesis and Antimicrobial Activity of Pseudodesmin and Viscosinamide Cyclic Lipopeptides Produced by Pseudomonads Associated with the Cocoyam Rhizosphere. Microorganisms 2020, 8, 1079.
- Pedras, M.S.C.; Ismail, N.; Quail, J.W.; Boyetchko, S.M. Structure, chemistry, and biological activity of pseudophomins A and B, new cyclic lipodepsipeptides isolated from the biocontrol bacterium Pseudomonas fluorescens. Phytochemistry 2003, 62, 1105–1114.
- Zachow, C.; Tilcher, R.; Berg, G. Sugar Beet-Associated Bacterial and Fungal Communities Show a High Indigenous Antagonistic Potential Against Plant Pathogens. Microb. Ecol. 2007, 55, 119–129.
- Zachow, C.; Jahanshah, G.; de Bruijn, I.; Song, C.; Ianni, F.; Pataj, Z.; Gerhardt, H.; Pianet, I.; Lämmerhofer, M.; Berg, G.; et al. The Novel Lipopeptide Poaeamide of the Endophyte Pseudomonas poae RE*1-1-14 Is Involved in Pathogen Suppression and Root Colonization. Mol. Plant Microbe Interact. 2015, 28, 800–810.
- Cantore, P.L.; Lazzaroni, S.; Coraiola, M.; Serra, M.D.; Cafarchia, C.; Evidente, A.; Iacobellis, N.S. Biological Characterization of White Line-Inducing Principle (WLIP) Produced by Pseudomonas reactans NCPPB1311. Mol. Plant Microbe Interact. 2006, 19, 1113–1120.
- Andersen, J.B.; Koch, B.; Nielsen, T.H.; Sørensen, D.; Hansen, M.; Nybroe, O.; Christophersen, C.; Sørensen, J.; Molin, S.; Givskov, M. Surface motility in Pseudomonas sp. DSS73 is required for efficient biological containment of the root-pathogenic microfungi Rhizoctonia solani and Pythium ultimum. Microbiology 2003, 149, 37–46.
- Götze, S.; Herbst-Irmer, R.; Klapper, M.; Görls, H.; Schneider, K.R.A.; Barnett, R.; Burks, T.; Neu, U.; Stallforth, P. Structure, Biosynthesis, and Biological Activity of the Cyclic Lipopeptide Anikasin. ACS Chem. Biol. 2017, 12, 2498–2502.
- Klapper, M.; Götze, S.; Barnett, R.; Willing, K.; Stallforth, P. Bacterial Alkaloids Prevent Amoebal Predation. Angew. Chem.—Int. Ed. 2016, 55, 8944–8947.
- Nielsen, T.H.; Sørensen, J. Production of Cyclic Lipopeptides by Pseudomonas fluorescens Strains in Bulk Soil and in the Sugar Beet Rhizosphere. Appl. Environ. Microbiol. 2003, 69, 861–868.
- Sørensen, D.; Nielsen, T.H.; Sørensen, J.; Christophersen, C. Cyclic lipoundecapeptide lokisin from Pseudomonas sp. strain DSS41. Tetrahedron Lett. 2002, 43, 4421–4423.
- Hultberg, M.; Alsberg, T.; Khalil, S.; Alsanius, B. Suppression of disease in tomato infected by Pythium ultimum with a biosurfactant produced by Pseudomonas koreensis. Entomophaga 2009, 55, 435–444.
- Gu, Y.; Ma, Y.; Wang, J.; Xia, Z.; Wei, H. Genomic insights into a plant growth-promoting Pseudomonas koreensis strain with cyclic lipopeptide-mediated antifungal activity. MicrobiologyOpen 2020, 9, e1092.
- Schlusselhuber, M.; Godard, J.; Sebban, M.; Bernay, B.; Garon, D.; Seguin, V.; Oulyadi, H.; Desmasures, N. Characterization of Milkisin, a Novel Lipopeptide With Antimicrobial Properties Produced By Pseudomonas sp. UCMA 17988 Isolated From Bovine Raw Milk. Front. Microbiol. 2018, 9, 1030.
- Nielsen, T.; Thrane, C.; Christophersen, C.; Anthoni, U.; Sorensen, J. Structure, production characteristics and fungal antagonism of tensin—a new antifungal cyclic lipopeptide from Pseudomonas fluorescens strain 96.578. J. Appl. Microbiol. 2000, 89, 992–1001.
- Nguyen, D.D.; Melnik, A.V.; Koyama, N.; Lu, X.; Schorn, M.; Fang, J.; Aguinaldo, K.; Lincecum, T.L., Jr.; Ghequire, M.G.; Carrion, V.J.; et al. Indexing the Pseudomonas specialized metabolome enabled the discovery of poaeamide B and the bananamides. Nat. Microbiol. 2016, 2, 16197.
- Omoboye, O.O.; Geudens, N.; Duban, M.; Chevalier, M.; Flahaut, C.; Martins, J.C.; Leclère, V.; Oni, F.E.; Höfte, M. Pseudomonas sp. COW3 Produces New Bananamide-Type Cyclic Lipopeptides with Antimicrobial Activity against Pythium myriotylum and Pyricularia oryzae. Molecules 2019, 24, 4170.
- Jahanshah, G.; Yan, Q.; Gerhardt, H.; Pataj, Z.; Lämmerhofer, M.; Pianet, I.; Josten, M.; Sahl, H.-G.; Silby, M.W.; Loper, J.E.; et al. Discovery of the Cyclic Lipopeptide Gacamide A by Genome Mining and Repair of the Defective GacA Regulator in Pseudomonas fluorescens Pf0-1. J. Nat. Prod. 2019, 82, 301–308.
- Compeau, G.; Al-Achi, B.J.; Platsouka, E.; Levy, S.B. Survival of rifampin-resistant mutants of Pseudomonas fluorescens and Pseudomonas putida in soil systems. Appl. Environ. Microbiol. 1988, 54, 2432–2438.
- Haas, D.; Défago, G. Biological control of soil-borne pathogens by fluorescent pseudomonads. Nat. Rev. Genet. 2005, 3, 307–319.
- Ma, Z.; Geudens, N.; Kieu, N.P.; Sinnaeve, D.; Ongena, M.; Martins, J.C.; Höfte, M. Biosynthesis, Chemical Structure, and Structure-Activity Relationship of Orfamide Lipopeptides Produced by Pseudomonas protegens and Related Species. Front. Microbiol. 2016, 7, 382.
- Paulsen, I.T.; Press, C.M.; Ravel, J.; Kobayashi, D.Y.; Myers, G.S.A.; Mavrodi, D.V.; DeBoy, R.T.; Seshadri, R.; Ren, Q.; Madupu, R.; et al. Complete genome sequence of the plant commensal Pseudomonas fluorescens Pf-5. Nat. Biotechnol. 2005, 23, 873–878.
- Howell, C.R.; Stipanovic, R.D. Control of Rhizoctonia Solani on Cotton Seedlings with Pseudomonas fluorescens and with an Antibiotic Produced by the Bacterium by the Soil Tube Method Described Previously. Phytopathology 1979, 69, 480–482.
- D’Aes, J.; Kieu, N.P.; Leclere, V.; Tokarski, C.; Olorunleke, F.E.; De Maeyer, K.; Jacques, P.; Höfte, M.; Ongena, M. To settle or to move? The interplay between two classes of cyclic lipopeptides in the biocontrol strain Pseudomonas CMR12a. Environ. Microbiol. 2014, 16, 2282–2300.
- Van Der Voort, M.; Meijer, H.J.G.; Schmidt, Y.; Watrous, J.; Dekkers, E.; Mendes, R.; Dorrestein, P.C.; Gross, H.; Raaijmakers, J.M. Genome mining and metabolic profiling of the rhizosphere bacterium Pseudomonas sp. SH-C52 for antimicrobial compounds. Front. Microbiol. 2015, 6, 693.
- Girard, L.; Höfte, M.; De Mot, R. Lipopeptide families at the interface between pathogenic and beneficial Pseudomonas-plant interactions. Crit. Rev. Microbiol. 2020, 46, 397–419.
- Berry, C.L.; Brassinga, A.K.C.; Donald, L.J.; Fernando, W.G.D.; Loewen, P.C.; De Kievit, T.R. Chemical and biological characterization of sclerosin, an antifungal lipopeptide. Can. J. Microbiol. 2012, 58, 1027–1034.
- Zhao, H.; Liu, Y.-P.; Zhang, L.-Q. In silico and Genetic Analyses of Cyclic Lipopeptide Synthetic Gene Clusters in Pseudomonas sp. 11K1. Front. Microbiol. 2019, 10, 544.
- Li, W.; Rokni-Zadeh, H.; De Vleeschouwer, M.; Ghequire, M.G.K.; Sinnaeve, D.; Xie, G.-L.; Rozenski, J.; Madder, A.; Martins, J.C.; De Mot, R. The Antimicrobial Compound Xantholysin Defines a New Group of Pseudomonas Cyclic Lipopeptides. PLoS ONE 2013, 8, e62946.
- Wu, L.; Xiao, W.; Chen, G.; Song, D.; Khaskheli, M.A.; Li, P.; Zhang, S.; Feng, G. Identification of Pseudomonas mosselii BS011 gene clusters required for suppression of Rice Blast Fungus Magnaporthe oryzae. J. Biotechnol. 2018, 282, 1–9.
- Kruijt, M.; Tran, H.; Raaijmakers, J.M. Functional, genetic and chemical characterization of biosurfactants produced by plant growth-promoting Pseudomonas putida 267. J. Appl. Microbiol. 2009, 107, 546–556.
- Vallet-Gely, I.; Novikov, A.; Augusto, L.; Liehl, P.; Bolbach, G.; Péchy-Tarr, M.; Cosson, P.; Keel, C.; Caroff, M.; Lemaitre, B. Association of Hemolytic Activity of Pseudomonas entomophila, a Versatile Soil Bacterium, with Cyclic Lipopeptide Production. Appl. Environ. Microbiol. 2010, 76, 910–921.
- Rokni-Zadeh, H.; Li, W.; Sánchez-Rodríguez, A.; Sinnaeve, D.; Rozenski, J.; Martins, J.C.; De Mot, R. Genetic and Functional Characterization of Cyclic Lipopeptide White-Line-Inducing Principle (WLIP) Production by Rice Rhizosphere Isolate Pseudomonas putida RW10S2. Appl. Environ. Microbiol. 2012, 78, 4826–4834.
- Omoboye, O.O.; Oni, F.E.; Batool, H.; Yimer, H.Z.; De Mot, R.; Höfte, M. Pseudomonas Cyclic Lipopeptides Suppress the Rice Blast Fungus Magnaporthe oryzae by Induced Resistance and Direct Antagonism. Front. Plant Sci. 2019, 10, 901.
- Rokni-Zadeh, H.; Li, W.; Sánchez-Rodríguez, A.; Sinnaeve, D.; Rozenski, J.; Martins, J.C.; De Mot, R. Genetic and Functional Characterization of Cyclic Lipopeptide White-Line-Inducing Principle (WLIP) Production by Rice Rhizosphere Isolate Pseudomonas putida RW10S2. Appl. Environ. Microbiol. 2012, 78, 4826–4834. [Google Scholar] [CrossRef] [PubMed]
- Omoboye, O.O.; Oni, F.E.; Batool, H.; Yimer, H.Z.; De Mot, R.; Höfte, M. Pseudomonas Cyclic Lipopeptides Suppress the Rice Blast Fungus Magnaporthe oryzae by Induced Resistance and Direct Antagonism. Front. Plant Sci. 2019, 10, 901. [Google Scholar] [CrossRef] [PubMed]
- Mehnaz, S.; Saleem, R.S.Z.; Yameen, B.; Pianet, I.; Schnakenburg, G.; Pietraszkiewicz, H.; Valeriote, F.; Josten, M.; Sahl, H.-G.; Franzblau, S.G.; et al. Lahorenoic Acids A–C, ortho-Dialkyl-Substituted Aromatic Acids from the Biocontrol Strain Pseudomonas aurantiaca PB-St2. J. Nat. Prod. 2013, 76, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Thrane, C.; Nielsen, T.H.; Nielsen, M.N.; Sörensen, J.; Olsson, S. Viscosinamide-Producing Pseudomonas fluorescens DR54 Exerts a Biocontrol effect on Pythium ultimum in Sugar Beet Rhizosphere. Design 2000, 33, 139–146. [Google Scholar] [CrossRef]
- Thrane, C.; Nielsen, M.N.; Rensen, J.S.; Olsson, S. Pseudomonas fluorescens DR54 Reduces Sclerotia Formation, Biomass Development, and Disease Incidence of Rhizoctonia solani Causing Damping-Off in Sugar Beet. Microb. Ecol. 2001, 42, 438–445. [Google Scholar] [CrossRef]
- Tran, H.; Ficke, A.; Asiimwe, T.; Höfte, M.; Raaijmakers, J.M. Role of the cyclic lipopeptide massetolide A in biological control of Phytophthora infestans and in colonization of tomato plants by Pseudomonas fluorescens. New Phytol. 2007, 175, 731–742. [Google Scholar] [CrossRef]
- Van De Mortel, J.E.; De Vos, R.C.; Dekkers, E.; Pineda, A.; Guillod, L.; Bouwmeester, K.; Van Loon, J.J.A.; Dicke, M.; Raaijmakers, J.M. Metabolic and Transcriptomic Changes Induced in Arabidopsis by the Rhizobacterium Pseudomonas fluorescens SS101. Plant Physiol. 2012, 160, 2173–2188. [Google Scholar] [CrossRef]
- D’Aes, J.; Hua, G.K.H.; De Maeyer, K.; Pannecoucque, J.; Forrez, I.; Ongena, M.; Dietrich, L.; Thomashow, L.S.; Mavrodi, D.; Höfte, M. Biological Control of Rhizoctonia Root Rot on Bean by Phenazine- and Cyclic Lipopeptide-Producing Pseudomonas CMR12a. Phytopathology 2011, 101, 996–1004. [Google Scholar] [CrossRef] [PubMed]
- Olorunleke, F.E.; Hua, G.K.H.; Kieu, N.P.; Ma, Z.; Höfte, M. Interplay between orfamides, sessilins and phenazines in the control of Rhizoctonia diseases by Pseudomonas sp. CMR12a. Environ. Microbiol. Rep. 2015, 7, 774–781. [Google Scholar] [CrossRef]
- Ma, Z.; Hua, G.K.H.; Ongena, M.; Höfte, M. Role of phenazines and cyclic lipopeptides produced by Pseudomonas sp. CMR12a in induced systemic resistance on rice and bean. Environ. Microbiol. Rep. 2016, 8, 896–904. [Google Scholar] [CrossRef] [PubMed]
- Oni, F.E.; Olorunleke, O.F.; Höfte, M. Phenazines and cyclic lipopeptides produced by Pseudomonas sp. CMR12a are involved in the biological control of Pythium myriotylum on cocoyam (Xanthosoma sagittifolium). Biol. Control. 2018, 129, 109–114. [Google Scholar] [CrossRef]
- Ma, Z.; Ongena, M.; Höfte, M. The cyclic lipopeptide orfamide induces systemic resistance in rice to Cochliobolus miyabeanus but not to Magnaporthe oryzae. Plant Cell Rep. 2017, 36, 1731–1746. [Google Scholar] [CrossRef] [PubMed]
- Gross, H.; Stockwell, V.O.; Henkels, M.D.; Nowak-Thompson, B.; Loper, J.E.; Gerwick, W.H. The Genomisotopic Approach: A Systematic Method to Isolate Products of Orphan Biosynthetic Gene Clusters. Chem. Biol. 2007, 14, 53–63. [Google Scholar] [CrossRef]
- Le, C.; Kruijt, M.; Raaijmakers, J. Involvement of phenazines and lipopeptides in interactions between Pseudomonas species and Sclerotium rolfsii, causal agent of stem rot disease on groundnut. J. Appl. Microbiol. 2011, 112, 390–403. [Google Scholar] [CrossRef] [PubMed]
- Mendes, R.; Kruijt, M.; de Bruijn, I.; Dekkers, E.; Van Der Voort, M.; Schneider, J.H.; Piceno, Y.M.; DeSantis, T.Z.; Andersen, G.L.; Bakker, P.A.; et al. Deciphering the Rhizosphere Microbiome for Disease-Suppressive Bacteria. Science 2011, 332, 1097–1100. [Google Scholar] [CrossRef]
- Lavermicocca, P.; Ballio, A.; Camoni, L.; Di Giorgio, D.; Marchiafava, C. Studies on the Effect of Syringomycin and Syringopeptins on the Functions of Plant Mitochondria. In Pseudomonas Syringae Pathovars and Related Pathogens. Developments in Plant Pathology; Rudolph, K., Burr, T.J., Mansfield, J.W., Stead, D., Vivian, A., von Kietzell, J., Eds.; Springer: Dordrecht, The Netherlands, 1997; pp. 198–201. [Google Scholar]
- Bull, C.T.; Stack, J.P.; Smilanick, J.L. Pseudomonas syringae Strains ESC-10 and ESC-11 Survive in Wounds on Citrus and Control Green and Blue Molds of Citrus. Biol. Control 1997, 8, 81–88. [Google Scholar] [CrossRef]
- Hildebrand, P.D.; Braun, P.G.; McRae, K.B.; Lu, X. Role of the biosurfactant viscosin in broccoli head rot caused by a pectolytic strain of Pseudomonas fluorescens. Can. J. Plant Pathol. 1998, 20, 296–303. [Google Scholar] [CrossRef]
- Gerard, J.; Lloyd, R.; Barsby, T.; Haden, P.; Kelly, M.T.; Andersen, R.J. Massetolides A−H, Antimycobacterial Cyclic Depsipeptides Produced by Two Pseudomonads Isolated from Marine Habitats. J. Nat. Prod. 1997, 60, 223–229. [Google Scholar] [CrossRef]
- De Leij, F.A.A.M.; Sutton, E.J.; Whipps, J.M.; Fenlon, J.S.; Lynch, J.M. Impact of Field Release of Genetically Modified Pseudomonas fluorescens on Indigenous Microbial Populations of Wheat. Appl. Environ. Microbiol. 1995, 61. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, T.H.; Christophersen, C.; Anthoni, U.; Sorensen, J. Viscosinamide, a new cyclic depsipeptide with surfactant and antifungal properties produced by Pseudomonas fluorescens DR54. J. Appl. Microbiol. 1999, 87, 80–90. [Google Scholar] [CrossRef] [PubMed]
- Sinnaeve, D.; Michaux, C.; Van Hemel, J.; Vandenkerckhove, J.; Peys, E.; Borremans, F.A.; Sas, B.; Wouters, J.; Martins, J.C. Structure and X-ray conformation of pseudodesmins A and B, two new cyclic lipodepsipeptides from Pseudomonas bacteria. Tetrahedron 2009, 65, 4173–4181. [Google Scholar] [CrossRef]
- Mazzola, M.; Zhao, X.; Cohen, M.F.; Raaijmakers, J.M. Cyclic Lipopeptide Surfactant Production by Pseudomonas fluorescens SS101 Is Not Required for Suppression of Complex Pythium spp. Populations. Phytopathology 2007, 97, 1348–1355. [Google Scholar] [CrossRef] [PubMed]
- Han, F.; Mortishire-Smith, R.J.; Rainey, P.B.; Williams, D.H. Structure of the white-line-inducing principle isolated from Pseudomonas reactans. Acta Crystallogr. Sect. C Cryst. Struct. Commun. 1992, 48, 1965–1968. [Google Scholar] [CrossRef]
- Soler-Rivas, C.; Arpin, N.; Olivier, J.M.; Wichers, H. WLIP, a lipodepsipeptide of Pseudomonas “reactans”, as inhibitor of the symptoms of the brown blotch disease of Agaricus bisporus. J. Appl. Microbiol. 1999, 86, 635–641. [Google Scholar] [CrossRef]
- Zarvandi, S.; Bahrami, T.; Pauwels, B.; Asgharzadeh, A.; Hosseini-Mazinani, M.; Salari, F.; Girard, L.; De Mot, R.; Rokni-Zadeh, H. Draft Genome Sequence of Cyclic Lipopeptide Producer Pseudomonas sp. Strain SWRI103, Isolated from Wheat Rhizosphere. Microbiol. Resour. Announc. 2020, 9, e00538-20. [Google Scholar] [CrossRef]
- Jang, J.Y.; Yang, S.Y.; Kim, Y.C.; Lee, C.W.; Park, M.S.; Kim, J.C.; Kim, I.S. Identification of Orfamide A as an Insecticidal Metabolite Produced by Pseudomonas protegens F6. J. Agric. Food Chem. 2013, 61, 6786–6791. [Google Scholar] [CrossRef]
- Takeuchi, K.; Noda, N.; Someya, N. Complete Genome Sequence of the Biocontrol Strain Pseudomonas protegens Cab57 Discovered in Japan Reveals Strain-Specific Diversity of This Species. PLoS ONE 2014, 9, e93683. [Google Scholar] [CrossRef] [PubMed]
- Loper, J.E.; Henkels, M.D.; Rangel, L.I.; Olcott, M.H.; Walker, F.L.; Bond, K.L.; Kidarsa, T.A.; Hesse, C.N.; Sneh, B.; Stockwell, V.O.; et al. Rhizoxin analogs, orfamide A and chitinase production contribute to the toxicity of Pseudomonas protegens strain Pf-5 to Drosophila melanogaster. Environ. Microbiol. 2016, 18, 3509–3521. [Google Scholar] [CrossRef]
- Biessy, A.; Novinscak, A.; Blom, J.; Thomashow, L.S.; Cazorla, F.M.; Josic, D.; Filion, M. Diversity of Phytobeneficial Traits Revealed by Whole-Genome Analysis of Worldwide-Isolated Phenazine-Producing Pseudomonas spp. Environ. Microbiol. 2019, 21, 437–455. [Google Scholar]
- Geudens, N.; Nasir, M.N.; Crowet, J.-M.; Raaijmakers, J.M.; Fehér, K.; Coenye, T.; Martins, J.C.; Lins, L.; Sinnaeve, D.; Deleu, M. Membrane Interactions of Natural Cyclic Lipodepsipeptides of the Viscosin Group. Biochim. Biophys. Acta 2016, 1859, 331–339. [Google Scholar] [CrossRef]
- Watrous, J.; Roach, P.; Alexandrov, T.; Heath, B.S.; Yang, J.Y.; Kersten, R.D.; van der Voort, M.; Pogliano, K.; Gross, H.; Raaijmakers, J.; et al. Mass spectral molecular networking of living microbial colonies. Proc. Natl. Acad. Sci. USA 2012, 109, E1743–E1752. [Google Scholar] [CrossRef] [PubMed]
- Reder-Christ, K.; Schmidt, Y.; Dörr, M.; Sahl, H.-G.; Josten, M.; Raaijmakers, J.M.; Gross, H.; Bendas, G. Model membrane studies for characterization of different antibiotic activities of lipopeptides from Pseudomonas. Biochim. Biophys. Acta 2011, 1818, 566–573. [Google Scholar] [CrossRef]
- Eschmidt, R.; Eköberl, M.; Emostafa, A.; Ramadan, E.M.; Emonschein, M.; Jensen, K.B.; Ebauer, R.; Eberg, G. Effects of bacterial inoculants on the indigenous microbiome and secondary metabolites of chamomile plants. Front. Microbiol. 2014, 5, 64. [Google Scholar] [CrossRef]
- Vassilev, V.; Lavermicocca, P.; DI Giorgio, D.; Iacobellis, N.S. Production of syringomycins and syringopeptins by Pseudomonas syringae pv. atrofaciens . Plant Pathol. 1996, 45, 316–322. [Google Scholar] [CrossRef]
- Emanuele, M.; Scaloni, A.; Lavermicocca, P.; Jacobellis, N.; Camoni, L.; Di Giorgio, D.; Pucci, P.; Paci, M.; Segre, A.; Ballio, A. Corceptins, new bioactive lipodepsipeptides from cultures of Pseudomonas corrugata. FEBS Lett. 1998, 433, 317–320. [Google Scholar] [CrossRef]
- Quibod, I.L.; Grande, G.; Oreiro, E.G.; Borja, F.N.; Dossa, G.S.; Mauleon, R.; Cruz, C.V.; Oliva, R. Rice-Infecting Pseudomonas Genomes Are Highly Accessorized and Harbor Multiple Putative Virulence Mechanisms to Cause Sheath Brown Rot. PLoS ONE 2015, 10, e0139256. [Google Scholar] [CrossRef]
- Ballio, A.; Bossa, F.; Camoni, L.; Di Giorgio, D.; Flamand, M.-C.; Maraite, H.; Nitti, G.; Pucci, P.; Scaloni, A. Structure of fuscopeptins, phytotoxic metabolites of Pseudomonas fuscovaginae. FEBS Lett. 1996, 381, 213–216. [Google Scholar] [CrossRef]
- Flamand, M.-C.; Pelsser, S.; Ewbank, E.; Maraite, H. Production of syringotoxin and other bioactive peptides by Pseudomonas fuscovaginae. Physiol. Mol. Plant Pathol. 1996, 48, 217–231. [Google Scholar] [CrossRef]
- Patel, H.K.; Da Silva, D.P.; Devescovi, G.; Maraite, H.; Paszkiewicz, K.; Studholme, D.J.; Venturi, V. Draft Genome Sequence of Pseudomonas fuscovaginae, a Broad-Host-Range Pathogen of Plants. J. Bacteriol. 2012, 194, 2765–2766. [Google Scholar] [CrossRef] [PubMed]
- Bender, C.L.; Alarcón-Chaidez, F.; Gross, D.C. Pseudomonas syringae Phytotoxins: Mode of Action, Regulation, and Biosynthesis by Peptide and Polyketide Synthetases. Microbiol. Mol. Biol. Rev. 1999, 63, 266–292. [Google Scholar] [CrossRef]
- Lopes, L.D.; Weisberg, A.J.; Davis, E.W., 2nd; Varize, C.d.S.; Silva, M.d.C.P.E.; Chang, J.H.; Loper, J.E.; Andreote, F.D. Genomic and metabolic differences between Pseudomonas putida populations inhabiting sugarcane rhizosphere or bulk soil. PLoS ONE 2019, 14, e0223269. [Google Scholar] [CrossRef] [PubMed]
- Keshavarz-Tohid, V.; Vacheron, J.; Dubost, A.; Prigent-Combaret, C.; Taheri, P.; Tarighi, S.; Taghavi, S.M.; Moënne-Loccoz, Y.; Muller, D. Genomic, phylogenetic and catabolic re-assessment of the Pseudomonas putida clade supports the delineation of Pseudomonas alloputida sp. nov., Pseudomonas inefficax sp. nov., Pseudomonas persica sp. nov., and Pseudomonas shirazica sp. nov. Syst. Appl. Microbiol. 2019, 42, 468–480. [Google Scholar] [CrossRef] [PubMed]
- Girard, L.; Geudens, N.; Pauwels, B.; Höfte, M.; Martins, J.C.; De Mot, R. Transporter Gene-mediated Typing for Detection and Genome Mining of Lipopeptide-producing Pseudomonas. Appl. Environ. Microbiol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Kuiper, I.; Lagendijk, E.L.; Pickford, R.; Derrick, J.; Lamers, G.E.M.; Thomas-Oates, J.E.; Lugtenberg, B.J.J.; Bloemberg, G.V. Characterization of two Pseudomonas putida lipopeptide biosurfactants, putisolvin I and II, which inhibit biofilm formation and break down existing biofilms. Mol. Microbiol. 2003, 51, 97–113. [Google Scholar] [CrossRef] [PubMed]
- Meziane, H.; Van der Sluis, I.; van Loon, L.C.; Höfte, M.; Bakker, P.A.H.M. Determinants of Pseudomonas putida WCS358 Involved in Inducing. Mol. Plant Pathol. 2005, 6, 177–185. [Google Scholar] [CrossRef]
- Berendsen, R.L.; Van Verk, M.C.; Stringlis, I.A.; Zamioudis, C.; Tommassen, J.; Pieterse, C.M.J.; Bakker, P.A.H.M. Unearthing the genomes of plant-beneficial Pseudomonas model strains WCS358, WCS374 and WCS417. BMC Genom. 2015, 16, 1–23. [Google Scholar] [CrossRef]
- Vodovar, N.; Vallenet, D.; Cruveiller, S.; Rouy, Z.; Barbe, V.; Acosta, C.; Cattolico, L.; Jubin, C.; Lajus, A.; Segurens, B.; et al. Complete genome sequence of the entomopathogenic and metabolically versatile soil bacterium Pseudomonas entomophila. Nat. Biotechnol. 2006, 24, 673–679. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.M.; Bakker, P.A.H.M. Induced Systemic Resistance by Beneficial Microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef]
- Schellenberger, R.; Touchard, M.; Clément, C.; Baillieul, F.; Cordelier, S.; Crouzet, J.; Dorey, S. Apoplastic invasion patterns triggering plant immunity: Plasma membrane sensing at the frontline. Mol. Plant Pathol. 2019, 20, 1602–1616. [Google Scholar] [CrossRef]
- Pel, M.J.C.; Pieterse, C.M.J. Microbial Recognition and Evasion of Host Immunity. J. Exp. Bot. 2012, 64, 1237–1248. [Google Scholar] [CrossRef]
- Behzadi, P.; García-Perdomo, H.A.; Karpiński, T.M. Toll-Like Receptors: General Molecular and Structural Biology. J. Immunol. Res. 2021, 2021, 1–21. [Google Scholar] [CrossRef]
- Medzhitov, R. Toll-like Receptors and Innate Immunity. Nat. Rev. Immunol. 2001, 1, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Grandellis, C.; Garavaglia, B.S.; Gottig, N.; Lonez, C.; Ruysschaert, J.-M.; Ottado, J. DOTAP, a lipidic transfection reagent, triggers Arabidopsis plant defense responses. Planta 2018, 249, 469–480. [Google Scholar] [CrossRef]
- Piasecka, A.; Jedrzejczak-Rey, N.; Bednarek, P. Secondary metabolites in plant innate immunity: Conserved function of divergent chemicals. New Phytol. 2015, 206, 948–964. [Google Scholar] [CrossRef]
- Zipfel, C.; Oldroyd, G.E.D. Plant signalling in symbiosis and immunity. Nature 2017, 543, 328–336. [Google Scholar] [CrossRef]
- Stringlis, I.A.; Zhang, H.; Pieterse, C.M.J.; Bolton, M.D.; de Jonge, R. Microbial small molecules—weapons of plant subversion. Nat. Prod. Rep. 2018, 35, 410–433. [Google Scholar] [CrossRef] [PubMed]
- Yu, K.; Yu, K.; Pieterse, C.M.; Pieterse, C.M.; Bakker, P.A.; Bakker, P.A.; Berendsen, R.L.; Berendsen, R.L.; Yu, K.; Yu, K.; et al. Beneficial microbes going underground of root immunity. Plant Cell Environ. 2019, 42, 2860–2870. [Google Scholar] [CrossRef] [PubMed]
- Spence, C.; Alff, E.; Johnson, C.; Ramos, C.; Donofrio, N.; Sundaresan, V.; Bais, H. Natural rice rhizospheric microbes suppress rice blast infections. BMC Plant Biol. 2014, 14, 130. [Google Scholar] [CrossRef]
- Brodey, C.L.; Rainey, P.B.; Tester, M.; Johnstone, K. Bacterial Blotch Disease of the Cultivated Mushroom Os Caused By an Iron Channel Forming Lipidesipsipeptide Toxin.Pdf. Mol. Plant. Microbe Interact. 1991, 4, 407–411. [Google Scholar] [CrossRef]
- Kozlova, O.V.; Egorov, S.Y.; Kupriyanova-Ashina, F.G.; Rid, N.; El’-Registan, G.I. Analysis of the Ca2+ response of mycelial fungi to external effects by the recombinant aequorin method. Microbiology 2004, 73, 629–634. [Google Scholar] [CrossRef]
- Aiyar, P.; Schaeme, D.; García-Altares, M.; Flores, D.C.; Dathe, H.; Hertweck, C.; Sasso, S.; Mittag, M. Antagonistic bacteria disrupt calcium homeostasis and immobilize algal cells. Nat. Commun. 2017, 8, 1–13. [Google Scholar] [CrossRef]
- Rainey, P.; Brodey, C.L.; Johnstone, K. Biological properties and spectrum of activity of tolaasin, a lipodepsipeptide toxin produced by the mushroom pathogen Pseudomonas tolaasii. Physiol. Mol. Plant Pathol. 1991, 39, 57–70. [Google Scholar] [CrossRef]
- Steigenberger, J.; Verleysen, Y.; Geudens, N.; Martins, J.C.; Heerklotz, H. The Optimal Lipid Chain Length of a Membrane-Permeabilizing Lipopeptide Results From the Balance of Membrane Partitioning and Local Damage. Front. Microbiol. 2021, 12, 1–14. [Google Scholar] [CrossRef]
- Leveau, J.H.J.; Preston, G. Bacterial mycophagy: Definition and diagnosis of a unique bacterial–fungal interaction. New Phytol. 2008, 177, 859–876. [Google Scholar] [CrossRef]
- Wong, W.C.; Preece, T.F. Identification of Pseudomonas tolaasi: The White Line in Agar and Mushroom Tissue Block Rapid Pitting Tests. J. Appl. Bacteriol. 1979, 47, 401–407. [Google Scholar] [CrossRef]
- De Vleeschouwer, M.; Van Kersavond, T.; Verleysen, Y.; Sinnaeve, D.; Coenye, T.; Martins, J.C.; Madder, A. Identification of the Molecular Determinants Involved in Antimicrobial Activity of Pseudodesmin A, a Cyclic Lipopeptide From the Viscosin Group. Front. Microbiol. 2020, 11, 646. [Google Scholar] [CrossRef]
- De Bruijn, I.; de Kock, M.J.D.; de Waard, P.; van Beek, T.A.; Raaijmakers, J.M. Massetolide A Biosynthesis in Pseudomonas fluorescens. J. Bacteriol. 2008, 190, 2777–2789. [Google Scholar] [CrossRef] [PubMed]
- Saxon, E.B.; Jackson, R.W.; Bhumbra, S.; Smith, T.; Sockett, R.E. Bdellovibrio bacteriovorus HD100 guards against Pseudomonas tolaasii brown-blotch lesions on the surface of post-harvest Agaricus bisporus supermarket mushrooms. BMC Microbiol. 2014, 14, 163. [Google Scholar] [CrossRef]
- Jia, J.; Wang, X.; Deng, P.; Ma, L.; Baird, S.M.; Li, X.; Lu, S. Pseudomonas glycinae sp. nov. isolated from the soybean rhizosphere. MicrobiologyOpen 2020, 9, e1101. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.; Rha, E.; Kim, H.; Lee, S.-G. Complete Genome Sequence of the Soil Bacterium Pseudomonas kribbensis Strain 46-2 T. Microbiol. Resour. Announc. 2018, 7, e01161-18. [Google Scholar] [CrossRef] [PubMed]
- Pascual, J.; García-López, M.; Bills, G.; Genilloud, O. Pseudomonas granadensis sp. nov., a new bacterial species isolated from the Tejeda, Almijara and Alhama Natural Park, Granada, Spain. Int. J. Syst. Evol. Microbiol. 2015, 65, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Cautain, B.; de Pedro, N.; Schulz, C.; Pascual, J.; Sousa, T.D.S.; Martínez, I.G.; Pérez-Victoria, I.; Asensio, F.; González, I.; Bills, G.; et al. Identification of the Lipodepsipeptide MDN-0066, a Novel Inhibitor of VHL/HIF Pathway Produced by a New Pseudomonas Species. PLoS ONE 2015, 10, e0125221. [Google Scholar] [CrossRef]
- Berry, C.L.; Nandi, M.; Manuel, J.; Brassinga, A.K.C.; Fernando, W.D.; Loewen, P.C.; de Kievit, T.R. Characterization of the Pseudomonas sp. DF41 quorum sensing locus and its role in fungal antagonism. Biol. Control. 2014, 69, 82–89. [Google Scholar] [CrossRef]
- Grgurina, I.; Mariotti, F.; Fogliano, V.; Gallo, M.; Scaloni, A.; Iacobellis, N.S.; Cantore, P.L.; Mannina, L.; Castelli, V.V.A.; Greco, M.L.; et al. A new syringopeptin produced by bean strains of Pseudomonas syringae pv. syringae. Biochim. Biophys. Acta 2002, 1597, 81–89. [Google Scholar] [CrossRef]
- Ballio, A.; Barra, D.; Bossa, F.; Collina, A.; Grgurina, I.; Marino, G.; Moneti, G.; Paci, M.; Pucci, P.; Segre, A.; et al. Syringopeptins, new phytotoxic lipodepsipeptides of Pseudomonas syringae pv. syringae. FEBS Lett. 1991, 291, 109–112. [Google Scholar] [CrossRef]
- Ravindran, A.; Jalan, N.; Yuan, J.S.; Wang, N.; Gross, D.C. Comparative genomics of Pseudomonas syringae pv. syringae strains B301D and HS 191 and insights into intrapathovar traits associated with plant pathogenesis. MicrobiologyOpen 2015, 4, 553–573. [Google Scholar] [CrossRef]
- Berti, A.D.; Greve, N.J.; Christensen, Q.H.; Thomas, M.G. Identification of a Biosynthetic Gene Cluster and the Six Associated Lipopeptides Involved in Swarming Motility of Pseudomonas syringae pv. tomato DC3000. J. Bacteriol. 2007, 189, 6312–6323. [Google Scholar] [CrossRef]
- Ramkumar, G.; Lee, S.W.; Weon, H.-Y.; Kim, B.-Y.; Lee, Y.H. First report on the whole genome sequence of Pseudomonas cichoriistrain JBC1 and comparison with other Pseudomonas species. Plant Pathol. 2015, 64, 63–70. [Google Scholar] [CrossRef]
- Alam, K.; Islam, M.; Li, C.; Sultana, S.; Zhong, L.; Shen, Q.; Yu, G.; Hao, J.; Zhang, Y.; Li, R.; et al. Genome Mining of Pseudomonas Species: Diversity and Evolution of Metabolic and Biosynthetic Potential. Molecules 2021, 26, 7524. [Google Scholar] [CrossRef] [PubMed]
This entry is offline, you can click here to edit this entry!