Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Subjects:
Agriculture, Dairy & Animal Science
The Brachystelma genus (family: Apocynaceae) consists of geophytes that are traditionally utilised among rural communities, especially in East Africa, southern Africa, West Africa, and northern and western India. Apart from being used as a food source, they are indicated as treatment for ailments such as colds, chest pains, and wounds. This review provides a critical appraisal on the ethnobotanical uses, nutritional value, phytochemical profiles, and biological activities of Brachystelma species.
- Apocynaceae
- conservation
- ethnobotany
- food crop
- micropropagation
- phenolics
1. Distribution and Botanical Description of Brachystelma Species
1.1. Distribution of Brachystelma Species
In the Ceropegieae tribe, Brachystelma is regarded as the second largest genus [1,2]. It is distributed, in order of decreasing diversity, in sub-Saharan Africa, India, Southeast Asia, and Australia [8]. As indicated by Masinde [10], about 90% of Brachystelma species occur exclusively in sub-Saharan Africa. In South Africa, Brachystelma species occur across the country i.e., in all nine provinces. Relative to the 85 Brachystelma species in the South African Red List data, approximately 73% are endemic to South Africa (Figure 1; Supplementary Table S1). In India, Brachystelma species are still being discovered and identified [20,23,24], which is an indication of their expanding diversity.
Evident from their distribution, Brachystelma species are variable in habitat preference. Some are found on mountain slopes with up to 3000 mm of annual precipitation, whereas others are found among rocks in grasslands with up to 200 mm of annual precipitation [10,20,23]. However, a common factor is the requirement of well-drained soils [10]. The grassland habitat makes them difficult to locate among the grasses. Furthermore, their visibility is restricted because they are geophytic and generally appear above ground during the rainy season [6,25,26].
1.2. Overview of Botanical Aspects of Brachystelma Species
Morphologically, members of the Brachystelma genus are relatively uniform in terms of their vegetative parts (Figure 2A–C). They are considered small perennial geophytic herbs arising from depressed globose or discoid tubers with fibrous roots originating from the sides and the base [10,27]. The tubers range from 15 to 200 mm in diameter, and the tuber size is said to correlate to the age of the plant [9,10]. From the rootstock arises one or generally a cluster of deciduous stem (Figure 2B). Their leaves are simple opposite, with or without stalks, and may be extremely variable in shape, even on a single plant [4,10,15]. Unlike the majority of its milky sap-producing Apocynaceae members, Brachystelma leaves are known to produce a clear sap [10]. The leaves are always non-succulent, and usually finely haired [4]. The flowers are inconspicuous, especially in a field among other vegetation [4,15]. They are often flat to cylindrical, come in one to several, and are found together on the side of the stem or rarely terminal, with short to medium-length stalks.

Figure 2. Morphology of some Brachystelma R. Br. ex Sims species. (A) Flowers (left to right—Brachystelma gerrardii; Brachystelma pulchellum; Brachystelma pygmaeum subsp. flavidum; Brachystelma pygmaeum subsp. pygmaeum); (B) leaves (left to right—Brachystelma gerrardii; Brachystelma ngomense; Brachystelma pulchellum; Brachystelma pygmaeum subsp. flavidum); (C) tubers and roots (Brachystelma pygmaeum subsp. pygmaeum). © Adam Shuttleworth.
Their corolla is rotate to tubular, divided to halfway or further down the base with five lobes (star-like), and free at the tips and variously spreading, or united at the tips, forming a cage-like structure [4,15]. The flowers are generally dark (brown to maroon, or pinkish purple) and accompanied by a slightly unpleasant odour, which is the primary attraction of pollinators (true flies) [4,5,9,15].
For most Brachystelma species, the pollinia are usually ovoid and are more or less D- shaped, sub-erect to erect, and near or on top of the staminal column [8,10,15]. Generally, the fruits are erect, long, or short in fusiform pairs. Once dry, the fruit (now follicle) contains the seeds [3,9]. The seeds are dark brown or light black and the size range is 5–10 mm long by 1.5–7 mm wide [8]. One end of the seed has a coma comprising a tuft of fine, white, fluffy hairs up to 20 mm long, which allow for wind dispersal [10,15,27].
1.3. Taxonomy of Brachystelma Species
The importance of the accurate naming of plant species cannot be over-emphasised [22]. A few inconsistencies were evident in the naming of some of the Brachystelma species reported in the eligible literature. Some of the observations included the incorrect spelling and use of synonyms for a few Brachystelma species. This clearly supports the call to make reference to reference databases for the validation of plant names.
2. Ethnobotanical Applications of Brachystelma Species
Many Brachystelma species are known by diverse names among different ethnic groups, which is an indication of their value and importance. The tubers are more widely utilised than other plant parts such as the leaves, roots, and stem. This is an indication that the tubers are highly valued in ethnobotany given their extensive utilisation across many countries. The ethnobotanical uses of approximately 34 Brachystelma species have been documented in 13 countries (e.g., Australia, Botswana, Burkina Faso, India, Lesotho, Namibia, Nigeria, South Africa, Tanzania, and Thailand) across the world. Based on the number of citations, ranging from three to six mentions, Brachystelma barberiae, Brachystelma dinteri, Brachystelma circinnatum, and Brachystelma foetidum were identified as the most popular species. Even though the majority of the Brachystelma species common in Africa have diverse uses, limited applications were indicated for Brachystelma species (e.g., Brachystelma glabriflorum and Brachystelma kerrii) found in Asian countries such as Cambodia and Thailand [28,29,30].
In some cases, local uses were identified on a regional basis, such as for Brachystelma johnstonii in East Africa [10] and Brachystelma circinnatum in southern Africa [31,32,33,34,35,36]. Generally, the diverse uses recorded for the 34 Brachystelma species can be categorised into two major areas, as highlighted below.
2.1. Uses as Food and for Nutritional Needs
The underground parts (particularly the tubers) of many Brachystelma species are known to retain a large amount of water and nutrients, thereby making them highly desirable as a food source. The tubers of several Brachystelma species (e.g., Brachystelma burchellii, Brachystelma dinteri, Brachystelma schultzei, Brachystelma thunbergii, and Brachystelma tuberosum) are eaten raw or prepared by indigenous groups in Africa [10,34,37,38], Asia [17,27], and Australia [39]. This is particularly important when there is limited food supply or during hunting expeditions. In East Africa (Kambaland) and South Africa, some Brachystelma species (e.g., Brachystelma gracile and Brachystelma johnstonii) are reported to be particularly enjoyed by livestock herders [10,15]. In Lesotho, the local name ‘Bohobe-ba-setsomi’ loosely translates to “bread of the hunters”, clearly denoting the value and importance of Brachystelma species [33]. Other food-related applications for Brachystelma species include as vegetables, snacks, and sweet preserves.
Given the high water content, the tubers of Brachystelma species are often considered an effective resource for quenching thirst in dry areas where fresh water is scarce [15,36,40]. In Burkina Faso, roots and tubers of Brachystelma bingeri are commonly used as a thirst quencher among the nomadic and hunting populations located in Nayala province [40]. Masinde [10] indicated similar uses for Brachystelma simplex, Brachystelma rubellum, and Brachystelma plocamoides in East Africa. The moisture content for the tubers is often high, reaching 80–97% , which justifies their uses as thirst quenchers among many indigenous groups.
Some wild animals are believed to harvest and feed on Brachystelma tubers [10]. Brachystelma species are explored for their nutritional value by animals. On the basis of causal observations [41], animals such as porcupines, mole rats, and blesmols, as well as baboons, are often the major competitors for all underground storage organs, including the deeply buried ones. Based on available anecdotal data, there are no reports of any Brachystelma species being poisonous to humans or livestock [10].
2.2. Applications as Herbal Medicine
Some Brachystelma species have also been reported as medicinal herbs across several countries. The East African species Brachystelma buchananii and Brachystelma johnstonii are regarded as medicinal herbs with diverse healing properties [10]. For instance, the tuber of Brachystelma buchananii is chewed and thereafter, the paste is placed on a wound. In northern Uganda, the tuber of Brachystelma johnstonii is dried, ground, and taken to relieve chest pains [10]. Evidence of the medicinal uses has been reported for Brachystelma foetidum in Lesotho [37,42] and Brachystelma togoense in Nigeria [43,44]. In India, Brachystelma edulis is used to exert therapeutic benefits for a wide range of health conditions, including headaches and stomach-aches [45,46]. The medicinal effect of other species, including Brachystelma kerrii, has been recorded in Cambodia and Thailand [29,30].
Based on the number of mentions, Brachystelma edulis, Brachystelma naorojii, and Brachystelma togoense were identified as species with diverse medicinal applications . Consensus on the uses of medicinal plants is an indication of their potential therapeutic values [47]. Some degree of similarity in uses was demonstrated with the application of Brachystelma for managing respiratory-related conditions such as colds, coughs, and chest pain in three countries, including China, India, and Lesotho. However, different Brachystelma species were utilised for cold in these aforementioned countries. These include Brachystelma edule (China), Brachystelma naorojii (India), and Brachystelma foetidum (Lesotho).
3. Nutritional and Phytochemical Aspects
3.1. Nutritional Composition of Brachystelma Species
Despite the relatively large number of Brachystelma species with an ethobotanical record as wild food, only Brachystelma edulis and Brachystelma naorojii currently have empirical data on their nutritional content. Nutritional analysis for wild edible plants that serve as food is essential to justify their incorporation into the diet [12,41,54,55]. Based on the study conducted by Deshmukh and Rathod [46], the tubers of Brachystelma edulis could be a good supplement for nutrients such as protein, fibre, and carbohydrates. The recorded amount for these aforementioned nutrients is within the range for common foods, especially vegetables [56]. Particularly, the level of carbohydrates among Brachystelma species was comparable to African leafy vegetables such as Bidens pilosa, Chenopodium album, and Portulaca oleracea [57]. The fibre content in Brachystelma edulis (8%) tubers compared favourably to Ceropegia hirsuta (9.1%), a closely-related species [46]. A similar trend was observed with regards to the crude protein level between these two aforementioned plants.
For some of the content (e.g., ash and moisture), comparable levels were present in the leaves and tubers, which is an indication of plant-part substitution with respect to Brachystelma edulis [27,45,46] and Brachystelma naorojii [58]. However, in terms of the dry matter (% DW), the tubers were generally higher than the leaves for both Brachystelma edulis [45,46] and Brachystelma naorojii [58]. Existing evidence demonstrated some degree of variation in parameters, such as ash and dry matter between Brachystelma edulis and Brachystelma naorojii,
Mineral analysis of Brachystelma edulis tubers revealed the presence of 9 elements with high concentrations of potassium, phosphorus, and magnesium [46]. These minerals are essential for maintaining good health and well-being [56,57,59]. Based on the concentrations of these minerals found in Brachystelma edulis, it certainly competes favourably with many indigenous vegetables (e.g., Amaranthus spinosus, Bidens pilosa, Chenopodium album, and Momordica involucrata), which are often sourced by local communities to meet their daily requirements [55]. However, the absence of nutritional and mineral content for the majority of Brachystelma species remains a major gap requiring empirical data to support their overwhelming use as a food source among different ethnic groups. Particularly, the potential development of novel food products from indigenous plants, including Brachystelma species, will be driven by the availability of basic and in-depth data on their nutritional value and sensory attributes [60].
3.2. Phytochemical Profile of Brachystelma Species
Among the Ceropegieae, Brachystelma have not received much attention in terms of the quantification of their phytochemicals. Currently, the phytochemical profiles of five Brachystelma species have been investigated using different analytical techniques, including qualitative and quantitative tests. Most studies search first for alkaloids, as they are known among the Apocynaceae to possess a range of potent pharmacological activities, including antibacterial, anticancer, and antimalarial activities [13,61]. Phytochemical analysis of Brachystelma edulis, based on qualitative tests, indicated the presence of alkaloids, cardiac glycosides, phenols, saponins, and tannins . Similarly, Brachystelma bingeri contains saponins, triterpenes, and sterols [62]. Furthermore, a spectrophotometric test of Brachystelma bingeri revealed an estimated 1.7 mg EGA/g (equivalent of gallic acid) of total phenolics, whereas the total flavonoids and tannins were only found in trace quantities.
Brachystelma pulchellum and Brachystelma pygmaeum also demonstrated the presence of phenolic acids, including both hydroxybenzoic and hydroxcinnamic acids [63]. The phenolic acid profile of in vitro grown Brachystelma pulchellum and Brachystelma pygmaeum showed similarities in their phenolic profiles, but quantities varied among species. Given the importance of hydroxybenzoic and hydroxcinnamic acids [64,65], their presence in these aforementioned Brachystelma species provide evidence on the potential health benefits associated with their consumption.
Some Brachystelma species, including Brachystelma ngomense and Brachystelma pulchellum, have been observed to have a fibrous stem covered by bark that appears red-violet in colour. Even though not well characterised in Brachystelma (or in Apocynaceae), red bark has generally been reported to be characteristic of chromoalkaloids (betacyanins) located in the cellular vacuoles of sub-epidermal tissue [66,67,68]. Certainly for Brachystelma species, this is an area that still requires stringent research [8].
4. Biological Activities of Brachystelma Species Extracts and Isolated Compounds
Based on indigenous knowledge, several Brachystelma species are known for their therapeutic effects for different health conditions, including the treatment of coughs and colds, wounds, and bodily discomfort, and for enhancing fertility. The rich phytochemical pools in a few Brachystelma species have been established from the existing limited studies. The presence of therapeutic phytochemicals are generally known to be responsible for the biological effect of medicinal plants [69]. Despite insufficient data on the biological activities of Brachystelma species, the tubers and leaves of a few species have been evaluated for antioxidant and acetylcholinesterase inhibitory effects as well as acute toxicity. In terms of the antioxidant effects, four Brachystelma species (Brachystelma bingeri, Brachystelma edulis, Brachystelma pulchellum, and Brachystelma pygmaeum) have been assessed using diverse assays, including ABTS, DPPH, FRAP, and ORAC methods. The use of different antioxidant assays is beneficial, as it has the potential to effectively reveal all the mechanisms associated with the evaluated plant [70]. However, no noteworthy antioxidant effects were observed among the evaluated Brachystelma species. Likewise, Brachystelma bingeri had moderate (50%) acetylcholine esterase (AChE) inhibition at a tested concentration of 100 μg/mL [62]. Using an in vivo model, a methanol extract of Brachystelma binger tubers caused no mortality to the tested mice and was considered safe at a concentration of up to 3000 mg/kg body weight [62].
The biological activity of isolated compounds from the aerial parts of Brachystelma togoense has predominantly targeted different microbes (e.g., Escherichia coli, Staphylococcus aureus, Streptococcus typhi, Staphylococcus pneumoniae, and Candida albicans) as a means of establishing their antimicrobial properties. As revealed by Ekalu et al. [43], phaeophytin A had the most promising antimicrobial effect (MIC = 0.09 mg/mL) against Candida albicans. Furthermore, the MIC value ranged from 0.18 to 0.37 mg/mL against the four tested bacterial strains (Escherichia coli, Staphylococcus aureus, Streptococcus typhi, and Staphylococcus pneumoniae). Relative to the recommended MIC (<100 μg/mL), the antimicrobial property of the majority of compounds isolated from Brachystelma togoense are classified as weak to moderate effects, thereby suggesting limited clinical significance [71].
5. Conservation Status of Brachystelma Species and Sustainability
5.1. Conservation Status of Brachystelma Species
In South Africa, many members (54%) of the genus Brachystelma remain of conservation concern (Figure 3; Supplementary Table S1). Furthermore, the threats on members of the Brachystelma genus have been reported in India [8]. Findings by Singh [17] revealed that Brachystelma laevigatum and Brachystelma pauciflorum are currently vulnerable and endangered due to their use by local people as food and for their attractive foliage. According to Rajakullayiswamy et al. [20], Brachystelma volubile is currently indicated as being critically endangered. In India, the grasslands in which Brachystelma thrive are often considered “wasted and unproductive” when left in the hands of nature and thus left unprotected [16]. In addition, these grasslands are often encroached on for agriculture and fragmented by urbanisation and invasive species.
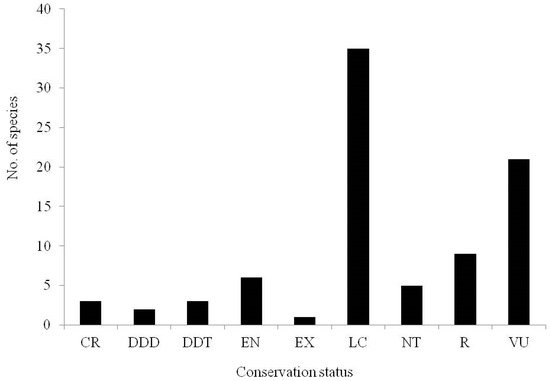
Figure 3. An overview of the conservation status of 85 Brachystelma species recorded in South Africa based on the South African National Biodiversity Institute (SANBI) Red List [21]. CR—Critically rare, DDD—Data deficient (insufficient information), DDT—Data deficient (taxonomically problematic), EN—Endangered, EX—Extinct, LC—Least concern, NT—Near threatened, R—Rare, VU—Vulnerable.
5.2. Propagation of Brachystelma Species
Natural cultivation of Brachystelma species has remained neglected and sparingly documented [2,8]. The most prevalent constraints, with natural propagation of species belonging to the subfamily Asclepiadoideae, are often attributed to the scarcity of pollinators together with low seed set and viability [75,76,77]. Propagation via conventional methods i.e., cuttings and seedlings, is also either absent or not well-documented among Brachystelma. Following pollination, the rate (%) of successful fertilisation varies among species, but is generally low when compared to many bee-pollinated species [15]. The pollination is affected by the deposit of pollinia on the flowers’ stigmatic surfaces fused within the top of the staminal column [8,15]. In terms of the arrangement of the male and female reproductive parts, they are combined into a small columnar structure within the base of the corolla, increasing the chances of self-pollination [15]. This inevitably affects the fertilisation due to the failure of producing viable seeds [76,78]. Upon successful fertilisation, fruit development is rapid but takes several weeks for the seeds to mature, and during this time, the fruit may be attacked by pests, thereby preventing seed maturation [15]. On this basis, efficient ex situ conservation measures are of importance for the future of the Brachystelma genus, especially when their nutritional and medicinal potential has not been fully assessed [8].
5.3. Micropropagation of Brachystelma Species
Globally, plant biotechnology, in particular, micropropagation, has remained valuable based on its beneficial role in ex situ conservation and the mass production of plants and plant-based resources [75,79,80,81,82,83]. Conventional methods alone are not sufficient to achieve optimum yield and production rates, especially in plants such as Brachystelma, which require long periods of time for bulb maturation [77,84]. The micropropagation process starts with the selection of appropriate explants till the successful establishment of the in vitro regenerants under ex vitro conditions [83]. The different stages need to be carefully implemented to achieve the overall goal of conserving the selected plant species.
Micropropagation attempts for four Brachystelma species have been conducted by different researchers (Table 6). Even though some degree of success was achieved for Brachystelma glabrum and Brachystelma ngomense [84,85], the regeneration frequencies for Brachystelma pulchellum and Brachystelma pygmaeum were quite low [84]. Following the application of thidiazuron (TDZ) in various combinations with three auxins (indole-3-acetic acid, IAA; indole-3-butyric acid, IBA; and naphthalene acetic acid, NAA), shoot proliferation of Brachystelma glabrum was significantly enhanced with the use of TDZ (1 mg/mL) and NAA (0.5 mg/mL) relative to the control and other treatments [85]. The vital role of cytokinins was evident with the diverse regeneration and proliferation responses recorded among the three Brachystelma species [84]. Particularly for shoot proliferation of Brachystelma ngomense (a Brachystelma species that is categorised as an endangered plant), the use of meta-topolin riboside (mTR, 25 μM) was the most effective (producing an estimated four in vitro shoots) of the three cytokinins tested. On the other hand, BA (25 μM) and iP (25 μM) were more suitable for the micropropagation of Brachystelma pygmaeum and Brachystelma pulchellum, respectively. Furthermore, various combinations (types and concentrations) of cytokinins and auxins were applied for the in vitro flowering and tuberisation of Brachystelma glabrum [85]. Given the importance of plant growth regulators (especially cytokinins and auxins) in micropropagation protocols [79,81,86,87], it is important to continuously explore how the different classes of plant growth regulators can be used to improve the regeneration of Brachystelma species.
This entry is adapted from the peer-reviewed paper 10.3390/horticulturae8020122
This entry is offline, you can click here to edit this entry!