It is an understatement that mating and DNA transfer are key events for living organisms. Among the traits needed to facilitate mating, cell adhesion between gametes is a universal requirement. Thus, there should be specific properties for the adhesion proteins involved in mating. Biochemical and biophysical studies have revealed structural information about mating adhesins, as well as their specificities and affinities, leading to some ideas about these specialized adhesion proteins. Single-cell force spectroscopy (SCFS) has added important findings. In SCFS, mating cells are brought into contact in an atomic force microscope (AFM), and the adhesive forces are monitored through the course of mating. The results have shown some remarkable characteristics of mating adhesins and add knowledge about the design and evolution of mating adhesins.
- atomic force microscopy
- cell–cell mating
- adhesion
- single-cell force spectroscopy
- yeasts
- bacteria
- conjugation
- gametes
1. Introduction
2. Characteristics of SCFS
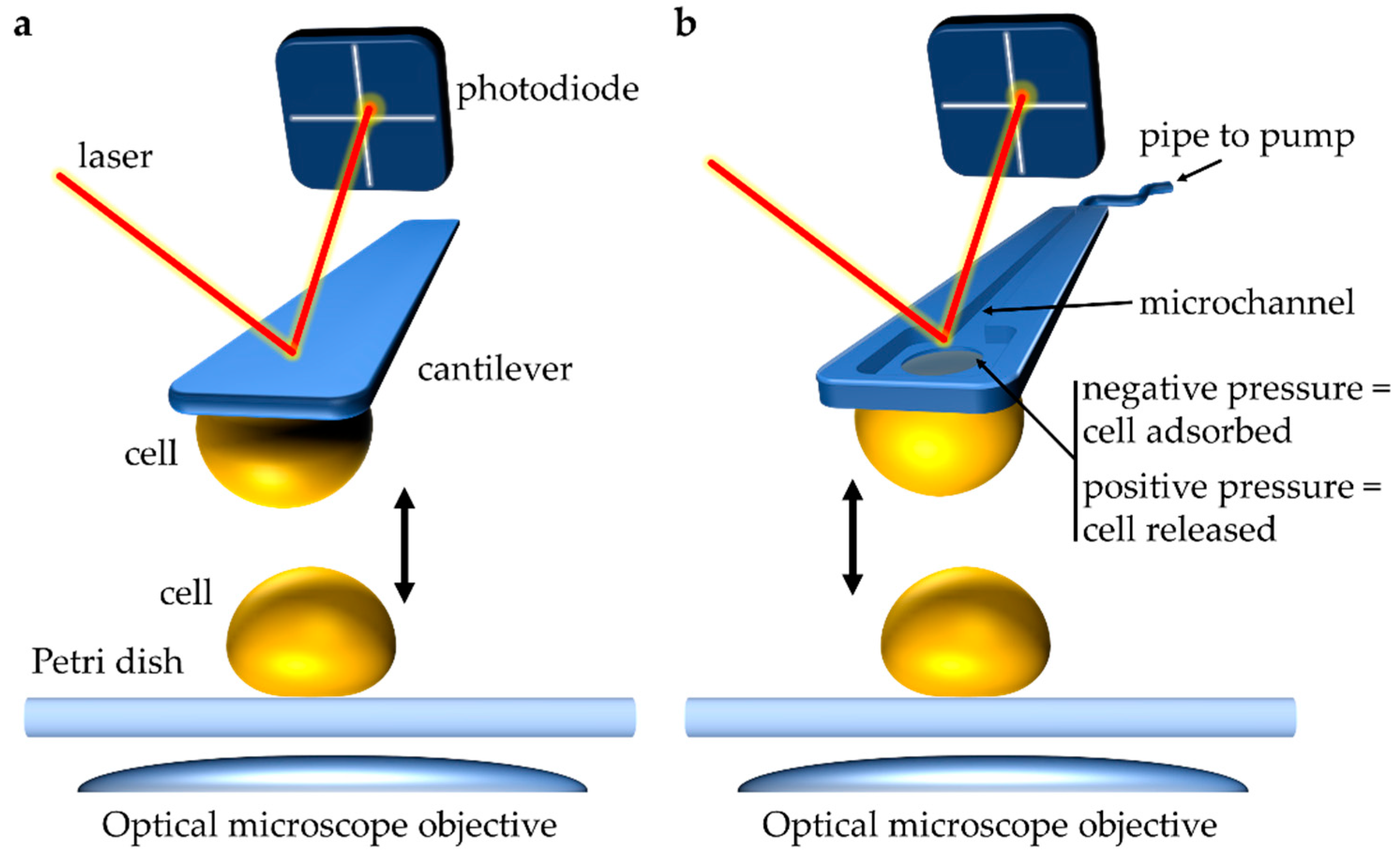 Figure 1. Atomic force microscopy (AFM) single-cell force spectroscopy (SCFS) to study cell–cell interactions during mating. (a) Conventional approach to performing single-cell SCFS experiments. A cell attached to a soft cantilever via (bio)chemical means (for example via poly-L-lysine coating) is brought into contact with a sample, like another cell. Interactive forces between the sample and the tip cause deflection of the cantilever. A laser beam focused on the cantilever and reflected onto a photodiode captures the deflection of the cantilever, which can be quantified as force (N). (b) In single-cell fluidic force microscopy (FluidFM) a micron-sized aperture connected to a microchannel within the cantilever is used to capture a cell through pressure differential. Connecting the cantilever through a microfluidics system to a pump allows controlling pressures at the apex. As such, the cantilever is approached toward a single cell, and a negative pressure is generated causing noninvasive adsorption of the cell to the cantilever. This cell probe can then be used to probe interactions with other single cells. Similarly, application of positive pressure can release the captured cell. This approach has aided in the study of yeast cell–cell interactions during mating.
Figure 1. Atomic force microscopy (AFM) single-cell force spectroscopy (SCFS) to study cell–cell interactions during mating. (a) Conventional approach to performing single-cell SCFS experiments. A cell attached to a soft cantilever via (bio)chemical means (for example via poly-L-lysine coating) is brought into contact with a sample, like another cell. Interactive forces between the sample and the tip cause deflection of the cantilever. A laser beam focused on the cantilever and reflected onto a photodiode captures the deflection of the cantilever, which can be quantified as force (N). (b) In single-cell fluidic force microscopy (FluidFM) a micron-sized aperture connected to a microchannel within the cantilever is used to capture a cell through pressure differential. Connecting the cantilever through a microfluidics system to a pump allows controlling pressures at the apex. As such, the cantilever is approached toward a single cell, and a negative pressure is generated causing noninvasive adsorption of the cell to the cantilever. This cell probe can then be used to probe interactions with other single cells. Similarly, application of positive pressure can release the captured cell. This approach has aided in the study of yeast cell–cell interactions during mating.2.1. SCFS Application to Studying Yeasts
2.1.1. Intercellular Adhesion
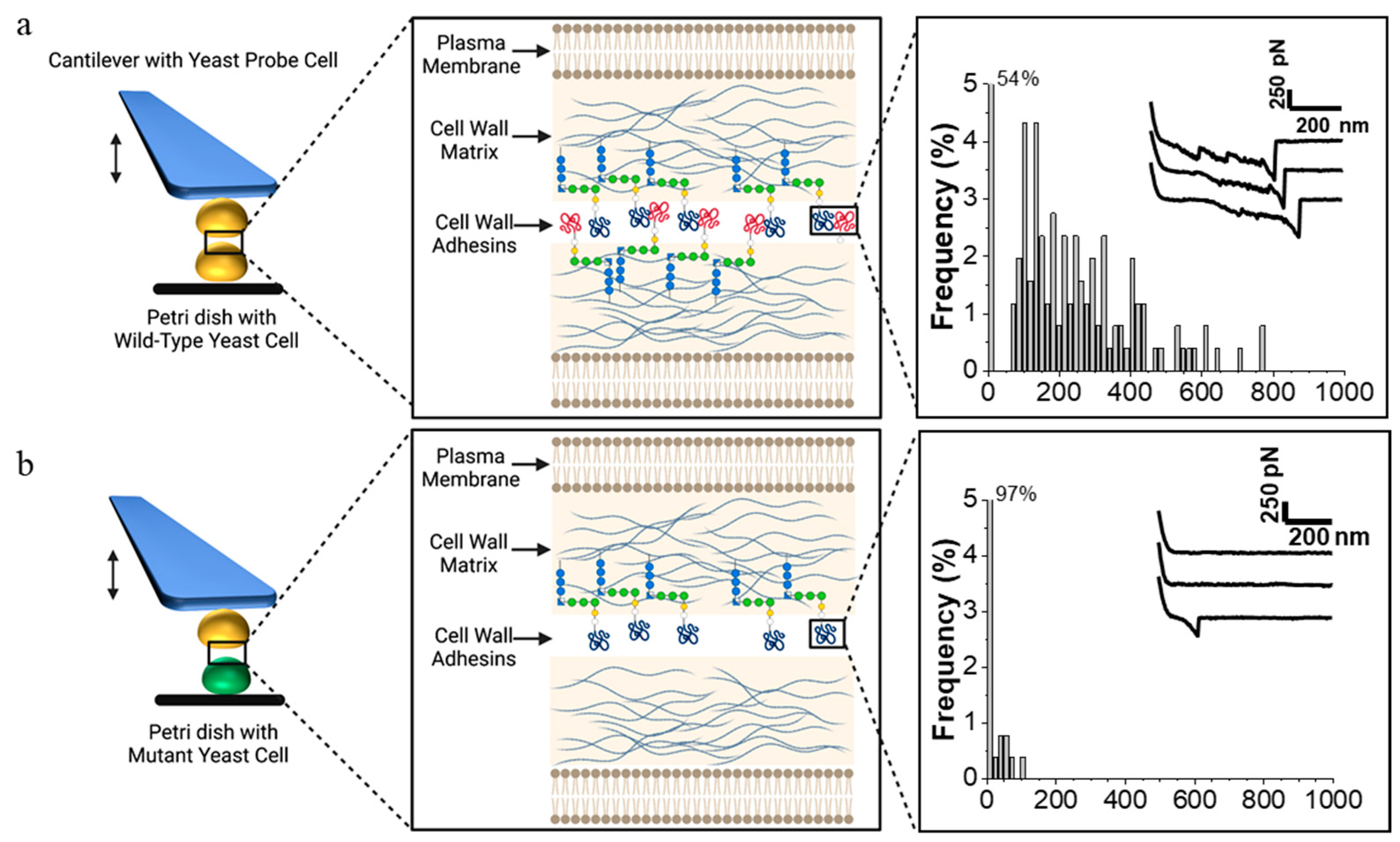 Figure 2. SCFS demonstration of the cellular role of a gene product in cell interaction. FluidFM-based SCFS allows for many different strains to be used as cell probes successively. Each cell probe can be brought into contact with many different cells in a petri dish. (a, left) A yeast cell probe attached to the cantilever interacts with a cell in the petri dish on the surface of the microscope stage through multiple cycles of contact and retraction of the probe. In the case of a mating experiment, the two cells are of opposite mating types. (middle) Details of cell surface showing cell membrane and cell wall, including blue glucan fibers and adhesins covalently crosslinked to the glucan through modified C-terminal GPI-anchors. (right) Histogram of rupture forces on successive adhesion events as the cells are brought into contact, and then separated; the inset shows force–distance curves for three representative contacts. (b) The situation when the cell on the microscope stage is a mutant cell, in this case, lacking a cell surface adhesin. This phenotype would result from mutation in the adhesin gene, in a gene required for cellular localization of the adhesin, or in a gene that regulates expression of these genes. In this case, the experiment yields infrequent cell–cell adhesions; thus, 97% of the F–D curves show no force needed for separation. The F–D plots are intended as illustrations only and are reprinted from Dehullu et al. [22].
Figure 2. SCFS demonstration of the cellular role of a gene product in cell interaction. FluidFM-based SCFS allows for many different strains to be used as cell probes successively. Each cell probe can be brought into contact with many different cells in a petri dish. (a, left) A yeast cell probe attached to the cantilever interacts with a cell in the petri dish on the surface of the microscope stage through multiple cycles of contact and retraction of the probe. In the case of a mating experiment, the two cells are of opposite mating types. (middle) Details of cell surface showing cell membrane and cell wall, including blue glucan fibers and adhesins covalently crosslinked to the glucan through modified C-terminal GPI-anchors. (right) Histogram of rupture forces on successive adhesion events as the cells are brought into contact, and then separated; the inset shows force–distance curves for three representative contacts. (b) The situation when the cell on the microscope stage is a mutant cell, in this case, lacking a cell surface adhesin. This phenotype would result from mutation in the adhesin gene, in a gene required for cellular localization of the adhesin, or in a gene that regulates expression of these genes. In this case, the experiment yields infrequent cell–cell adhesions; thus, 97% of the F–D curves show no force needed for separation. The F–D plots are intended as illustrations only and are reprinted from Dehullu et al. [22].2.1.2. Intercellular Adhesion during Mating
2.2. SCFS Application to Studying Bacteria
2.2.1. Bacterial Adhesion
2.2.2. Bacterial Adhesion in the Context of Mating
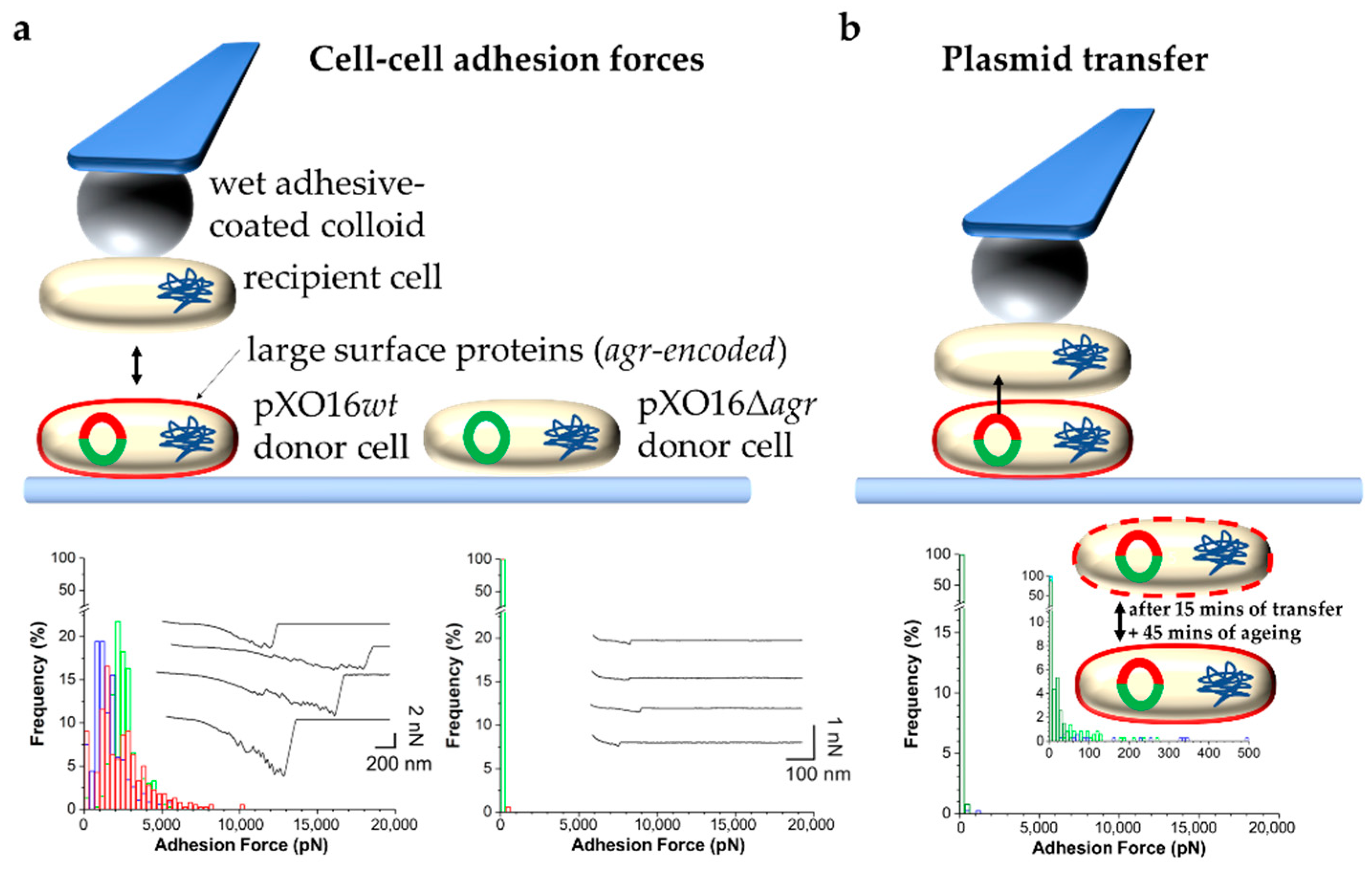 Figure 3. SCFS to probe and control bacterial conjugation. (a) Measuring forces in the adhesion of single pXO16wt or pXO16Δagr donor cells to single recipient cells. Top: Schematic of the approach. The pXO16 plasmid is represented by a red and green (pXO16wt) or green (pXO16Δagr) circle. The red part indicates the Agr region encoding large surface proteins (also indicated as a red outline around cells) that bind to recipient cells not expressing such proteins. Bottom: Histogram plots of the adhesion forces between recipient and donor cells carrying pXO16wt (left) pXO16Δagr (right). (b) Controlling plasmid transfer. Top: Schematic showing the approach consisting of (i) contact between recipient and donor cells for 15 min allowing DNA transfer, (ii) followed by retraction and rest for 45 min allowing maturation, i.e., proper expression and surface localization of agr-encoded proteins, and (iii) eventual measurements of cell–cell adhesion forces to evaluate changes in cell surface properties caused by plasmid transfer. Bottom: Histogram plots of the adhesion forces showing a loss of strong adhesion as a result of the presence of agr-encoded proteins on both cells. Using the resulting pXO16wt-cell probe to probe a naïve recipient cell restored strong adhesion as shown in the bottom left panel of (a). Figure adapted with permission from [35].
Figure 3. SCFS to probe and control bacterial conjugation. (a) Measuring forces in the adhesion of single pXO16wt or pXO16Δagr donor cells to single recipient cells. Top: Schematic of the approach. The pXO16 plasmid is represented by a red and green (pXO16wt) or green (pXO16Δagr) circle. The red part indicates the Agr region encoding large surface proteins (also indicated as a red outline around cells) that bind to recipient cells not expressing such proteins. Bottom: Histogram plots of the adhesion forces between recipient and donor cells carrying pXO16wt (left) pXO16Δagr (right). (b) Controlling plasmid transfer. Top: Schematic showing the approach consisting of (i) contact between recipient and donor cells for 15 min allowing DNA transfer, (ii) followed by retraction and rest for 45 min allowing maturation, i.e., proper expression and surface localization of agr-encoded proteins, and (iii) eventual measurements of cell–cell adhesion forces to evaluate changes in cell surface properties caused by plasmid transfer. Bottom: Histogram plots of the adhesion forces showing a loss of strong adhesion as a result of the presence of agr-encoded proteins on both cells. Using the resulting pXO16wt-cell probe to probe a naïve recipient cell restored strong adhesion as shown in the bottom left panel of (a). Figure adapted with permission from [35].3. Summary and Conclusions
This entry is adapted from the peer-reviewed paper 10.3390/ijms23031110