The soma, dendrites and axon of neurons may display calcium-dependent release of transmitters and peptides. Such release is named extrasynaptic for occurring in absence of synaptic structures. Emphasis is given to the somatic release of serotonin by the classical leech Retzius neuron, which has allowed detailed studies on the fine steps from excitation to exocytosis. Trains of action potentials induce transmembrane calcium entry through L-type channels. For action potential frequencies above 5 Hz, summation of calcium transients on individual action potentials activates the second calcium source: ryanodine receptors produce calcium-induced calcium release. The resulting calcium tsunami activates mitochondrial ATP synthesis to fuel transport of vesicles to the plasma membrane. Serotonin that is released maintains a large-scale exocytosis by activating the third calcium source: serotonin autoreceptors coupled to phospholipase C promote IP3 production. Activated IP3 receptors in peripheral endoplasmic reticulum release calcium that promotes vesicle fusion. The Swiss-clock workings of the machinery for somatic exocytosis has a striking disadvantage. The essential calcium-releasing endoplasmic reticulum near the plasma membrane hinders the vesicle transport, drastically reducing the thermodynamic efficiency of the ATP expenses and elevating the energy cost of release.
1. Introduction
Streams of action potentials or long depolarizations may evoke calcium-dependent exocytosis of transmitters and peptides from the soma, dendrites and axon of neurons in the absence of pre- and postsynaptic structures [
1,
2,
3,
4,
5,
6,
7,
8]. In contrast to the fast and localized release from synapses, extrasynaptic release is slow and diffuse, producing effects by means of volume transmission of signaling substances [
9]. Synaptic release contributes to rapid communication by neuronal circuits. By contrast, substances that are released extrasynaptically reach targets at variable distances to modulate the workings of populations of neurons, glia and blood vessels [
10,
11]. An extensive list of reviews on the effects of extrasynaptic exocytosis can be found in the literature [
8,
12,
13,
14,
15,
16,
17,
18,
19,
20].
All peptides and most classical low molecular weight transmitters have been shown to be released extrasynaptically from vesicles, either clear or often dense-core vesicles that rest distantly from the plasma membrane [
6]. Therefore, bursts of electrical activity or long depolarizations and calcium entry are needed to mobilize vesicles to the plasma membrane [
21,
22]. Calcium is not only essential for vesicle fusion, but has intermediate essential roles in the vesicle transport.
This review summarizes experimental evidence about the ways by which calcium flow through transmembrane and intracellular channels contributes in an orderly manner to induce vesicle transport and a large-scale exocytosis that lasts for hundreds of seconds. The backbone for the paper is experiments made on the soma of the classical large serotonergic Retzius neuron of the leech CNS, which allows direct experimentation to analyze the sequence of fine steps that couple electrical activity with exocytosis [
22]. Data supporting the generality of our results, obtained from different types of neurons, tissues and animals are mentioned along the text. For the moment it is useful to mention that electrical stimulation of a Retzius neuron with trains of 10 impulses at 20 Hz in an isolated ganglion activates the neuronal circuit that controls crawling locomotor behavior [
23]. During crawling, bursts of motoneuron firing produce alternate elongation of front body segments with shortening of rear segments [
24,
25]. The minute range latency to crawling is due first, to the series of calcium-dependent processes (the matter of this review), that take place between electrical activity and serotonin liberation. Unpublished experiments by Sanchez-Sugía and the author of this review show that the second part of the long latency to behavior reflects the arrival time of released serotonin to distant neuron targets that codify for crawling (Sanchez-Sugía C., and De-Miguel, F.F., in preparation).
2. Electrical Activity and Calcium Promote Vesicle Transport and Somatic Exocytosis
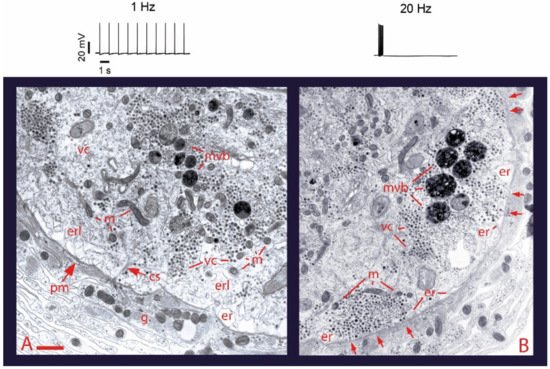
Ultrastructural analysis of the soma of Retzius neurons under different stimulation frequencies permits an initial understanding of the role of electrical stimulation and calcium on somatic exocytosis. The electron micrographs in
Figure 1 show the effects of electrical stimulation and calcium entry on somatic clusters of serotonin-containing dense core vesicles [
26]. The injection of brief depolarizing current pulses through a microelectrode to produce trains of action potentials at 1 Hz trains (
Figure 1), a frequency similar to that of spontaneous firing in serotonergic and other monoaminergic neurons [
27,
28,
29,
30], fails to promote any vesicle mobilization to the plasma membrane and exocytosis. However, increasing the frequency of the train to 20 Hz (
Figure 1B) produces ~50% of the vesicle clusters in electron micrographs to appear closely apposed to the plasma membrane [
26]. If a similar 20 Hz train is delivered in the presence of extracellular horseradish peroxidase, gold-coupled anti-peroxidase antibodies recognize peroxidase inside vesicles that underwent the exo-endocytosis cycle [
31]. Our laboratory has also found that after 20 Hz stimulation in the presence of extracellular magnesium to block transmembrane calcium entry, vesicles remain at resting positions (Del Pozo-Sánchez, V., Mendez, B. and De-Miguel, F.F., in preparation).
Figure 1. Electrical activity promotes vesicle transport to the plasma membrane. (
A) Electron micrograph of a soma fixed after 1 Hz stimulation (train on top). Vesicle clusters remain distant from the plasma membrane. (
B) After 20 Hz stimulation (top) vesicle clusters appear apposed to the plasma membrane (arrows). vc = vesicle clusters; mt = microtubules; pm = plasma membrane; m = mitochondria; er = endoplasmic reticulum; erl = internal endoplasmic reticulum layers; mvb = multivesicular bodies; g = glial cell processes. Scale bar = 1 µm. Obtained with permission from [
26].
3. Calcium Enters the Neuronal Soma through L-Type Channels
The first source of calcium for somatic exocytosis in Retzius neurons is the transmembrane flow through L-type (CaV1) channels [
32,
33]. L-type channels are well suited for somatic exocytosis in Retzius neurons, dopaminergic retinal neurons, serotonergic neuroepithelial neurons and in certain peptidergic neurons [
3,
32,
33,
34,
35], for their slow inactivation permits continuous calcium inflow during long depolarizations or trains of action potentials. In dendrites of magnocellular neurons releasing vasopressin and oxytocin and in other neuron types, N type (CaV2.2) channels contribute to exocytosis alone or in collaboration with L-type channels [
35,
36]. However, inactivation of N-type channels restricts their frequency-responses over long periods of electrical activity. In dendrites of magnocellular neurons, serotonergic neurons and dopamine ganglion neurons of the retina calcium elevations may result from activation of glutamate receptors, usually in the absence of local action potentials [
3,
37,
38].
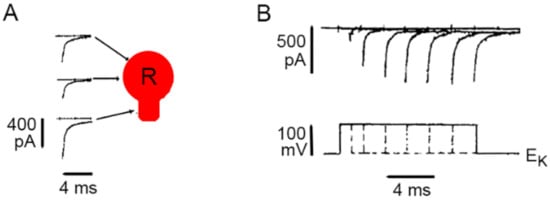
Evidence on the presence of L-type channels in the plasma membrane of the soma of Retzius neurons came first from loose patch clamp recordings of currents from small areas of plasma membrane [
39].
Figure 2 shows that the somatic calcium tail currents are smaller than those in the axon stump, a preferential region for the regeneration of neurites and synapse formation in culture [
39]. The kinetics of activation of somatic calcium currents was reconstructed by superposing isolated calcium tail currents recorded from a patch of membrane on increasingly larger depolarization voltage pulses. As shown in
Figure 2B, the calcium current inactivates slowly, as expected for L-type currents but not for calcium currents carried by other types of calcium channels.
Figure 2. Calcium currents though L-type channels in the soma membrane. (
A) Loose patch clamp recordings of calcium tail currents from small patches of membrane upon 10 ms depolarization. All recordings were obtained with the same pipette, thus making the amplitude of currents comparable for reflecting the density of active channels. (
B) Lack of inactivation of somatic calcium current indicating predominant activation of L-type channels. The activation kinetics were reconstructed by superimposing tail currents upon depolarizations with increasing durations. Potassium tail currents were prevented by clamping the voltage at the potassium equilibrium potential (E
K). Sodium tail currents were eliminated by reducing the sodium concentration in the pipette. Adapted with permission from the publisher of [
39].
4. High-Frequency Activation of L-Type Channels Promotes Somatic Exocytosis
Somatic exocytosis of serotonin can be evoked in Retzius cells in a frequency-dependent manner by trains of action potentials but not by individual action potentials [
32,
40]. Exocytosis on stimulation trains of ten impulses can be conveniently visualized from the formation of fluorescent spots of FM1-43 right inside the plasma membrane [
32]. Each spot forms upon the accumulation of dye inside a cluster of vesicles that undergo exo-endocytosis cycles. Although 1 Hz stimulation fails to evoke exocytosis, a number of fluorescent spots are produced by constitutive exocytosis. Such staining occurs equally well in the absence of stimulation, on 1 Hz stimulation, or on 20 Hz stimulation under experimental conditions that eliminate calcium entry [
32]. Above 5 Hz, the number of fluorescent spots increases with the stimulation frequency [
40].
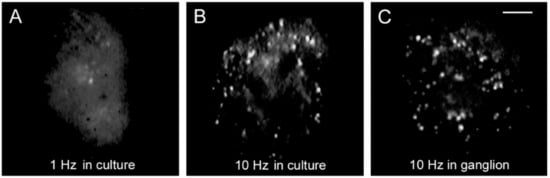
Figure 3 compares the FM1-43 staining patterns of cultured neurons, one stimulated with 1 Hz train (
Figure 3A) and another stimulated with 10 Hz train (
Figure 3B). The staining pattern produced in culture by 10 Hz stimulation reproduces that of neurons stimulated in the ganglion (
Figure 3C), therefore, the culture conditions provide an accurate bioassay to follow the sequence of events that led to serotonin exocytosis [
32]. A plateau in the amount of release at 20 Hz in Retzius neurons, in lobster serotonergic neurons and in mammalian peptidergic neurons [
40,
41,
42], suggests a generalized peak of efficiency for the coupling between electrical activity, calcium and exocytosis. For the reasons above, the experiments to be described next were carried out by stimulating neurons with 1 Hz or 20 Hz trains in combination with different experimental manipulations.
Figure 3. Somatic exocytosis as measured from FM1-43 fluorescence staining. (
A) The fluorescent FM1-43 spots after stimulation with a 1 Hz train correspond to constitutive exocytosis. (
B) Increase in the number of fluorescent spots after a 10 Hz train. (
C) Similar staining pattern in the soma of a neuron stimulated in the ganglion. Scale bar = 10 μm. Obtained with permission from [
32].
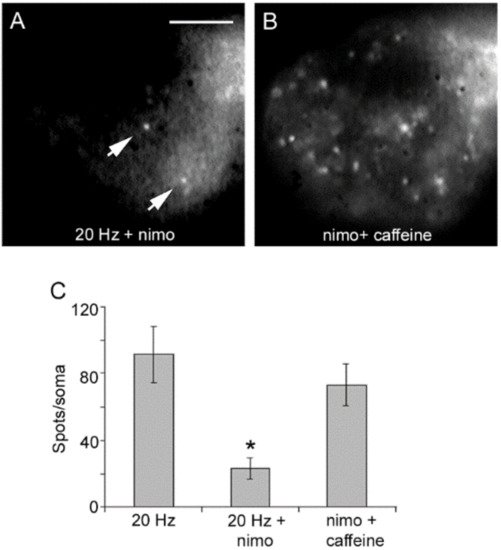
Blockade of L-type channels through the incubation of neurons with nimodipine abolishes exocytosis upon 20 Hz stimulation without affecting the constitutive exocytosis (
Figure 4A) [
32,
40]. Subsequent addition of caffeine to such same neurons to promote release of intracellular calcium through ryanodine receptors [
43,
44,
45,
46] in the absence of electrical stimulation produced a bulk of exocytosis, seen as an increased number of fluorescent spots (
Figure 4B). Such results clarified that calcium entry upon activation of L-type channels promotes both vesicle transport and exocytosis. Moreover, they suggested that calcium-induced calcium release contributes to link excitation and exocytosis.
Figure 4. Activation of L-type calcium channels mediate somatic exocytosis. (
A) Incubation with nimodipine (nimo) abolishes evoked but not constitutive exocytosis upon 20 Hz stimulation. The spots pointed by the arrow remained in B. (
B) Caffeine in the presence of nimodipine evokes vesicle transport and exocytosis in the absence of electrical stimulation. Images are selected areas of the soma. (
C) Exocytosis quantified as the number of fluorescent FM1-43 spots per soma. Bars are the S.E.M.; the * indicates a
p < 0.05 T test significance with respect to the 20 Hz and nimo+caffeine groups. The 20 Hz and nimo+caffeine groups were similar (
p > 0.05). Reproduced with permission from the publisher of [
32].
This entry is adapted from the peer-reviewed paper 10.3390/ijms23031495