Spelt wheat (Triticum aestivum L. ssp. spelta Thell.) is an ancient wheat that was widely cultivated in the past. This species derived from a cross between emmer wheat (T. turgidum spp. dicoccum Schrank em. Thell.) and Aegilops tauschii Coss. Its main origin was in the Fertile Crescent (Near East), with a secondary center of origin in Europe due to a second hybridization event between emmer and hexaploid wheat. This species has been neglected in most of Europe; however, the desire for more natural foods has driven a revival in interest. Iberian spelt is classified as a geographical group differing to the rest of European spelt.
- agrobiodiversity
- ancient wheat
- genetic resources
- neglected crops
1. Introduction
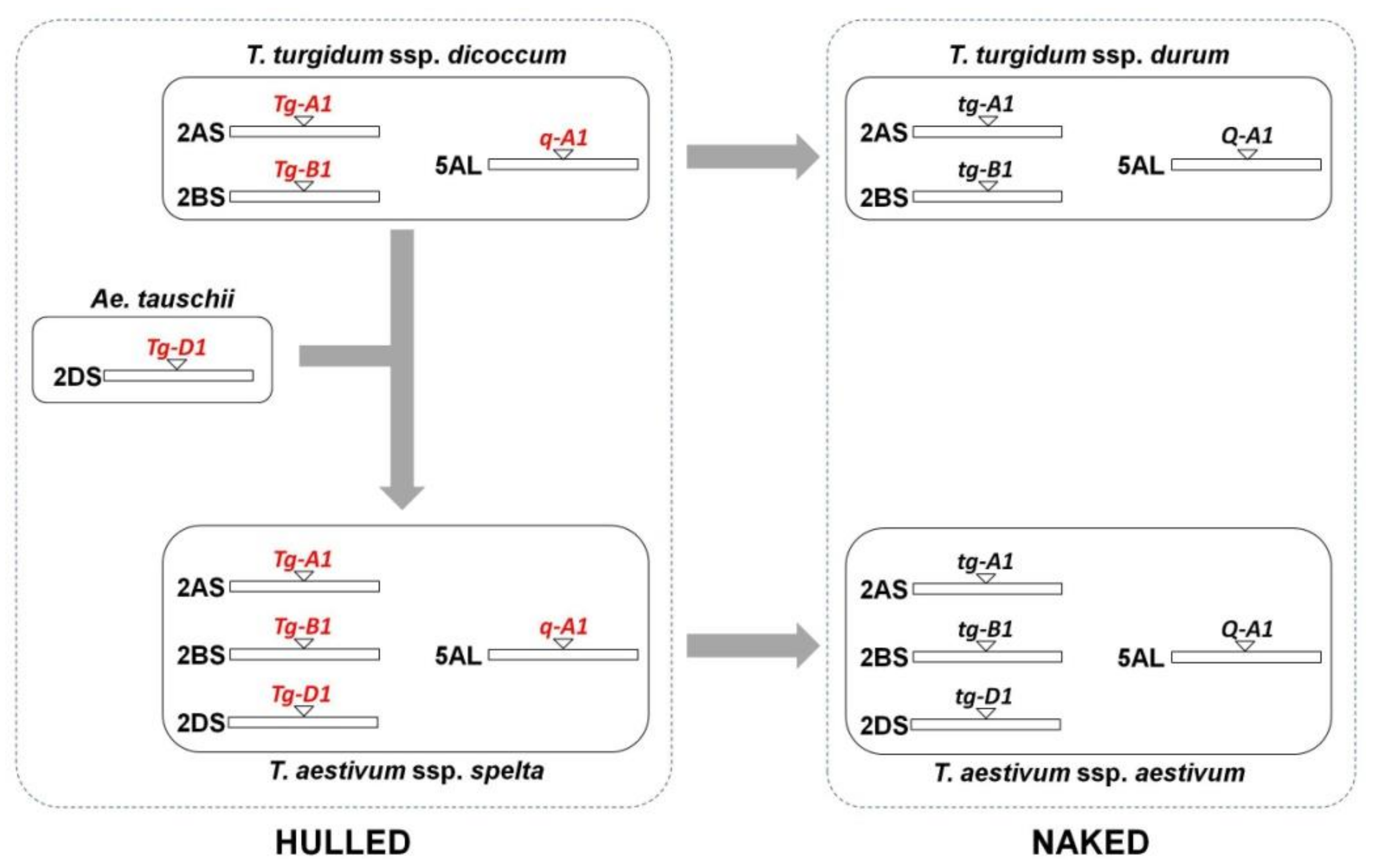
2. Recovery or Development of New Spelt Varieties


3. Perspectives
This entry is adapted from the peer-reviewed paper 10.3390/plants10122748
References
- Zohary, D.; Hopf, M. Domestication of Plants in the Old World: The Origin and Spread of Cultivated Plants in West Asia, Europe, and the Nile Valley; Oxford University Press: Oxford, UK, 2000.
- Harlan, J.R. Crops and Man; Crop Science Society of America and American Society of Agronomy: Madison, WI, USA, 1992.
- Giles, R.J.; Brown, T.A. GluDy allele variations in Aegilops tauschii and Triticum aestivum: Implications for the origins of hexaploid wheats. Theor. Appl. Genet. 2006, 112, 1563–1572.
- McFadden, E.S.; Sears, E.R. The artificial synthesis of Triticum spelta. Genetics 1945, 30, 14.
- Salamini, F.; Ozkan, H.; Brandolini, A.; Schafer-Pregl, R.; Martin, W. Genetics and geography of wild cereal domestication in the near east. Nat. Rev. Genet. 2002, 3, 429–441.
- Liu, Y.G.; Tsunewaki, K. Restriction-fragment-length-polymorphism (RFLP) analysis in wheat. 2. Linkage maps of the RFLP sites in common wheat. Jpn. J. Genet. 1991, 66, 617–633.
- Yan, Y.; Hsam, S.L.K.; Yu, J.Z.; Jiang, Y.; Ohtsuka, I.; Zeller, F.J. HMW and LMW glutenin alleles among putative tetraploid and hexaploid European spelt wheat (Triticum spelta L.) progenitors. Theor. Appl. Genet. 2003, 107, 1321–1330.
- Blatter, R.H.E.; Jacomet, S.; Schlumbaum, A. About the origin of European spelt (Triticum spelta L.): Allelic differentiation of the HMW Glutenin B1-1 and A1-2 subunit genes. Theor. Appl. Genet. 2004, 108, 360–367.
- Dedkova, O.S.; Badaeva, E.D.; Mitrofanova, O.P.; Zelenin, A.V.; Pukhalskiy, V.A. Analysis of intraspecific divergence of hexaploid wheat Triticum spelta L. by C-banding of chromosomes. Russ. J. Genet. 2004, 40, 1111–1126.
- Elía, M.; Moralejo, M.; Rodriguez-Quijano, M.; Molina-Cano, J.L. Spanish spelt: A separate gene pool within the spelt germplasm. Plant Breed. 2004, 123, 297–299.
- Muramatsu, M. Spike type in two cultivars of Triticum dicoccum with the spelta gene q compared with the Q-bearing variety liguliforme. Jpn. J. Breed. 1985, 35, 255–267.
- Kato, K.; Miura, H.; Sawada, S. QTL mapping of genes controlling ear emergence time and plant height on chromosome 5A of wheat. Theor. Appl. Genet. 1999, 98, 472–477.
- Kato, K.; Sonokawa, R.; Miura, H.; Sawada, S. Dwarfing effect associated with the threshability gene Q on wheat chromosome 5A. Plant Breed. 2003, 122, 489–492.
- Simons, K.J.; Fellers, J.P.; Trick, H.N.; Zhang, Z.; Tai, Y.-S.; Gill, B.S.; Faris, J.D. Molecular characterization of the major wheat domestication gene Q. Genetics 2006, 172, 547–555.
- Zhang, Z.C.; Belcram, H.; Gornicki, P.; Charles, M.; Just, J.; Huneau, C.; Magdelenat, G.; Couloux, A.; Samain, S.; Gill, B.S.; et al. Duplication and partitioning in evolution and function of homoeologous Q loci governing domestication characters in polyploid wheat. Proc. Natl. Acad. Sci. USA 2011, 108, 18737–18742.
- Debernardi, J.M.; Lin, H.; Chuck, G.; Faris, J.D.; Dubcovsky, J. microRNA172 plays a crucial role in wheat spike morphogenesis and grain threshability. Development 2017, 144, 1966–1975.
- Greenwood, J.R.; Finnegan, E.J.; Watanabe, N.; Trevaskis, B.; Swain, S.M. New alleles of the wheat domestication gene Q reveal multiple roles in growth and reproductive development. Development 2017, 144, 1959–1965.
- Kato, K.; Miura, H.; Akiyama, M.; Kuroshima, M.; Sawada, S. RFLP mapping of the three major genes, Vrn1, Q and B1, on the long arm of chromosome 5A of wheat. Euphytica 1998, 101, 91–95.
- Kerber, E.R.; Rowland, G.G. Origin of the free threshing character in hexaploid wheat. Can. J. Genet. Cytol. 1974, 16, 145–154.
- Simonetti, M.C.; Bellomo, M.P.; Laghetti, G.; Perrino, P.; Simeone, R.; Blanco, A. Quantitative trait loci influencing free-threshing habit in tetraploid wheats. Genet. Resour. Crop Evol. 1999, 46, 267–271.
- Dvorak, J.; Deal, K.R.; Luo, M.C.; You, F.M.; von Borstel, K.; Dehghani, H. The origin of spelt and free-threshing hexaploid wheat. J. Hered. 2012, 103, 426–441.
- Faris, J.; Zhang, Q.; Chao, S.; Zhang, Z.; Xu, S. Analysis of agronomic and domestication traits in a durum × cultivated emmer wheat population using a high-density single nucleotide polymorphism-based linkage map. Theor. Appl. Genet. 2014, 127, 2333–2348.
- Campbell, K.G. Spelt: Agronomy, genetics, and breeding. Plant Breed. Rev. 1997, 15, 187–214.
- Fischler, F. Commission Regulation (EC) No 1263/96 of 1 July 1996 supplementing the Annex to Regulation (EC) No 1107/96 on the registration of geographical indications and designations of origin under the procedure laid down in Article 17 of Regulation (EEC) No 2081/92. Off. J. Eur. Communities 1997, 163, 19–21.
- Caballero, L.; Martín, L.M.; Alvarez, J.B. Allelic variation of the HMW glutenin subunits in Spanish accessions of spelt wheat (Triticum aestivum ssp spelta L. em. Thell.). Theor. Appl. Genet. 2001, 103, 124–128.
- Schmid, J.; Winzeler, H. Genetic studies of crosses between common wheat (Triticum aestivum L.) and spelt (Triticum spelta L.). J. Genet. Breed. 1990, 44, 75–80.
- Winzeler, H.; Schmid, J.E.; Winzeler, M. Analysis of the yield potential and yield components of F1 and F2 hybrids of crosses between wheat (Triticum aestivum L.) and spelt (Triticum spelta L.). Euphytica 1994, 74, 211–218.
- Watson, A.; Ghosh, S.; Williams, M.J.; Cuddy, W.S.; Simmonds, J.; Rey, M.D.; Asyraf Md Hatta, M.; Hinchliffe, A.; Steed, A.; Reynolds, D.; et al. Speed breeding is a powerful tool to accelerate crop research and breeding. Nat. Plants 2018, 4, 23–29.
- Lantos, C.; Bóna, L.; Nagy, É.; Békés, F.; Pauk, J. Induction of in vitro androgenesis in anther and isolated microspore culture of different spelt wheat (Triticum spelta L.) genotypes. Plant Cell Tissue Organ Cult. 2018, 133, 385–393.
- Lantos, C.; Purgel, S.; Ács, K.; Langó, B.; Bóna, L.; Boda, K.; Békés, F.; Pauk, J. Utilization of in vitro anther culture in spelt wheat breeding. Plants 2019, 8, 436.
- Lantos, C.; Pauk, J. In vitro anther culture for doubled haploid plant production in spelt wheat. In Doubled Haploid Technology: Volume 1: General Topics, Alliaceae, Cereals; Segui-Simarro, J.M., Ed.; Springer: New York, NY, USA, 2021; pp. 257–266.
- Keller, M.; Karutz, C.; Schmid, J.E.; Stamp, P.; Winzeler, M.; Keller, B.; Messmer, M.M. Quantitative trait loci for lodging resistance in a segregating wheat × spelt population. Theor. Appl. Genet. 1999, 98, 1171–1182.
- Rachoń, L.; Bobryk-Mamczarz, A.; Kiełtyka-Dadasiewicz, A. Hulled wheat productivity and quality in modern agriculture against conventional wheat species. Agriculture 2020, 10, 275.
- Biel, W.; Jaroszewska, A.; Stankowski, S.; Sobolewska, M.; Kępińska-Pacelik, J. Comparison of yield, chemical composition and farinograph properties of common and ancient wheat grains. Eur. Food Res. Technol. 2021, 247, 1525–1538.
- Longin, C.F.H.; Ziegler, J.; Schweiggert, R.; Koehler, P.; Carle, R.; Würschum, T. Comparative study of hulled (einkorn, emmer, and spelt) and naked wheats (durum and bread wheat): Agronomic performance and quality traits. Crop Sci. 2016, 56, 302–311.
- Castillo, A.M.; Allue, S.; Costar, A.; Alvaro, F.; Valles, M.P. Doubled haploid production from Spanish and Central European spelt by anther culture. J. Agr. Sci. Technol. 2019, 21, 1313–1324.
- Peña-Chocarro, L. In situ conservation of hulled wheat species: The case of Spain. In Hulled Wheats; Padulosi, S., Hammer, K., Heller, J., Eds.; International Plant Genetic Resources Institute: Rome, Italy, 1996; pp. 129–146.
- Caballero, L.; Martín, L.M.; Alvarez, J.B. Agrobiodiversity of hulled wheats in Asturias (North of Spain). Genet. Resour. Crop Evol. 2007, 54, 267–277.
- Longin, C.F.H.; Würschum, T. Genetic variability, heritability and correlation among agronomic and disease resistance traits in a diversity panel and elite breeding material of spelt wheat. Plant Breed. 2014, 133, 459–464.
- Rapp, M.; Beck, H.; Gütler, H.; Heilig, W.; Starck, N.; Römer, P.; Cuendet, C.; Uhlig, F.; Kurz, H.; Würschum, T.; et al. Spelt: Agronomy, quality, and flavor of its breads from 30 varieties tested across multiple environments. Crop Sci. 2017, 57, 739–747.
- Packa, D.; Załuski, D.; Graban, Ł.; Lajszner, W. An evaluation of spelt crosses for breeding new varieties of spring spelt. Agronomy 2019, 9, 167.
- Ratajczak, K.; Sulewska, H.; Grażyna, S.; Matysik, P. Agronomic traits and grain quality of selected spelt wheat varieties versus common wheat. J. Crop Improv. 2020, 34, 654–675.
- Suchowilska, E.; Wiwart, M.; Krska, R.; Kandler, W. Do Triticum aestivum L. and Triticum spelta L. hybrids constitute a promising source material for quality breeding of new wheat varieties? Agronomy 2020, 10, 43.
- Takač, V.; Tóth, V.; Rakszegi, M.; Mikić, S.; Mirosavljević, M.; Kondić-Špika, A. Differences in processing quality traits, protein content and composition between spelt and bread wheat genotypes grown under conventional and organic production. Foods 2021, 10, 156.
- Callejo, M.J.; Vargas-Kostiuk, M.-E.; Ribeiro, M.; Rodríguez-Quijano, M. Triticum aestivum ssp. vulgare and ssp. spelta cultivars: 2. Bread-making optimisation. Eur. Food Res. Technol. 2019, 245, 1399–1408.
- Rodriguez-Quijano, M.; Nieto-Taladriz, M.T.; Carrillo, J.M. Polymorphism of waxy proteins in Iberian hexaploid wheats. Plant Breed. 1998, 117, 341–344.
- Alvarez, J.B.; Caballero, L.; Martín, L.M. Variability for morphological traits and high molecular weight glutenin subunits in Spanish spelt lines. Plant Genet. Resour.-Charact. Util. 2007, 5, 128–130.
- Rodriguez-Quijano, M.; Vazquez, J.F.; Carrillo, J.M. Variation of high molecular weight glutenin subunits in Spanish landraces of Triticum aestivum ssp. vulgare and ssp. spelta. J. Genet. Breed. 1990, 44, 121–126.
- Caballero, L.; Martín, L.M.; Alvarez, J.B. Genetic variability of the low-molecular-weight glutenin subunits in spelt wheat (Triticum aestivum ssp spelta L. em Thell.). Theor. Appl. Genet. 2004, 108, 914–919.
- Caballero, L.; Martín, L.M.; Alvarez, J.B. Intra- and interpopulation diversity for HMW glutenin subunits in Spanish spelt wheat. Genet. Resour. Crop Evol. 2004, 51, 175–181.
- Caballero, L.; Martín, L.M.; Alvarez, J.B. Variation and genetic diversity for gliadins in Spanish spelt wheat accessions. Genet. Resour. Crop Evol. 2004, 51, 679–686.
- Caballero, L.; Martín, L.M.; Alvarez, J.B. Relationships between the HMW- and LMW-glutenin subunits and SDS-sedimentation volume in spanish hulled wheat lines. Czech J. Genet. Plant Breed. 2008, 44, 114–117.
- Guzmán, C.; Caballero, L.; Moral, A.; Alvarez, J.B. Genetic variation for waxy proteins and amylose content in Spanish spelt wheat (Triticum spelta L.). Genet. Resour. Crop Evol. 2010, 57, 721–725.
- Guzmán, C.; Caballero, L.; Martín, L.M.; Alvarez, J.B. Waxy genes from spelt wheat: New alleles for modern wheat breeding and new phylogenetic inferences about the origin of this species. Ann. Bot. 2012, 110, 1161–1171.
- Guzmán, C.; Caballero, L.; Yamamori, M.; Alvarez, J.B. Molecular characterization of a new waxy allele with partial expression in spelt wheat. Planta 2012, 235, 1331–1339.