Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Subjects:
Agriculture, Dairy & Animal Science
The climate is one of the key elements impacting several cycles connected to soil and plant systems, as well as plant production, soil quality, and environmental quality. Due to heightened human activity, the rate of CO2 is rising in the atmosphere. Changing climatic conditions (such as temperature, CO2, and precipitation) influence plant nutrition in a range of ways, comprising mineralization, decomposition, leaching, and losing nutrients in the soil.
- climate change
- elevated CO2
- precipitation
- plant nutrition
1. Introduction
Human activity elevates the atmospheric levels of greenhouse gases (GHGs), causing the warming of the planet’s surface and a change in the climate [1][2][3]. Undoubtedly, climate change (CC) caused by global warming poses significant environmental risks [4]. Changes in CO2 levels in the atmosphere, temperature, and precipitation have a direct effect on agroecosystems (Figure 1), however, agroecosystems also account for almost one-third of overall GHG emissions, owing mostly to N fertilizers, livestock, and rice cultivation, and tropical deforestation [1][2]. There is much discussion over how varied the significance of the CC effects will be in different places throughout the world. The least developed and developing countries in the sub-tropics and tropics are more vulnerable to the adverse effects of CC. Crop productivity in low-income countries is projected to be negatively affected as a result of CC [2].

Figure 1. The effect of rising temperature and climate change on agroecosystems.
Carbon is an important element of the primary building block of all living beings for Earth’s life. It may be seen in a variety of forms, the most popular of which is plant biomass. Soil contains around 75% of the total C pool on the ground. As a result, soil is critical to sustaining a balanced global C cycle. Carbon dioxide content has increased by 30% in the atmosphere during the last 150 years [5]. The rising CO2 level of the atmosphere is a result of human activity and is linked to strong and perhaps catastrophic CC. Many solutions for dealing with soil misuse or degradation could help with CC adaptation [6]. One strategy proposed for reducing atmospheric CO2 is to enhance C storage in soils. As a result, there is an urgent need to manage soils and boost their capacity to retain more organic C [5][7]. Preventing and minimizing soil overuse, as well as encouraging sustainable land uses, could improve the fertility of the soil, enhance C stocks in soil and biomass, and help agricultural production and food security. As a result, soil nutrients are preserved and the impacts of climate change are mitigated [6].
2. Plant Nutrition under Climate Change
The plant needs some mineral nutrients for optimum growth and development. These nutrients are essential constituents of several macromolecules, which include nucleic acids, phospholipids, specific amino acids, and various co-enzymes, and they play an important role in plants’ cellular metabolism. Furthermore, they aid in chlorophyll production, redox processes, plasma membrane integrity, and cellosmotic potential [8]. The climate, as a key element of global ecosystems, has had a significant effect on the cycles and processes associated with humans, plants, and animals [9]. One of the most important elements influencing plant productivity is soil nutrition. Simultaneously, the availability of soil nutrients to plants varies widely among species [10]. Nutrient availability is vulnerable to CC. However, global warming may modify overall annual and seasonal nutrient availability and cycling [11]. In particular, variations in C, N, and P availability have serious impacts for plants because they are essential nutrients for plant development.
In addition, P has a major effect on water-use efficacy, regulating plant sensitivity to drought stress. These CCs, along with the significant spatial variability of soil nutrients and processes related to changes in habitat quality, result in a complicated scenario that influences soil microbial activity and hence nutrient availability for the plant. In addition, it is becoming increasingly obvious that variations in temperature or precipitation caused by CC change nutrient cycles and, as a result, plant nutrient availability [12]. Arndal et al. [13] reported that increased CO2, rising temperatures, and water stress are key variables that might alter future nutritional demand and availability. The growth of plants has been demonstrated to enhance and boost plant root biomass in many kinds of grassland when CO2 levels are increased. However, it is unclear whether plants will be able to exchange extra carbon for nutrients as a result of CC. Plant nutrient uptake will be controlled by compensative modifications under high CO2, and initiation of the mycorrhizal relationship is one of the potential mechanisms for improving nutrient uptake.
With CC, nutrient uptake will have to be preserved at current levels and also to be increased owing to the dispersion effect of more C assimilation to preserve nutrient concentrations in agricultural products. There is a functional equilibrium between shoot and root growth to improve a plant’s uptake of nutrients from the soil and atmosphere. The photosynthetic efficiency × leaf weight is correlated to the nutrient uptake × the root weight. If one of the previous four parameters is changed, the balance is changed unless some other parameter is also changed. In non-woody plants, increased CO2 in the atmosphere stimulates root and shoot development, with root growth exceeding shoot growth. Nevertheless, root growth is only encouraged if N is not a limiting factor [14]. However, the relationship between plant nutrition and climate change is extremely complicated because climatic circumstances impact all plant growth phases, including development, metabolism, physiology, and plant yields [15]. Agroecosystems capture water, light, CO2, and nutrients and use them to make various plant products such as proteins, carbohydrates, and starch. Solar radiation drives the total plant process, which is chemically converted via the photosynthesis process [16]. Globally, CC is a severe challenge that has simulated the curiosity of the world’s scientific community. The temperature rise caused by numerous human activities is the primary driving force behind CC. It has been observed that the average temperature of the surface of Earth has risen by about 0.9 °C since the late 19th century, with CO2 that caused by human activities being the primary cause of this rise [17]. Nonetheless, the CC affects temperature, water availability, and CO2, which affects the process of plant development and, ultimately, plants’ capacity to efficiently generate the abovementioned items that people require as food [16].
According to Dotaniya et al. [9], the overall impact of CC will decrease SOC pools, releasing more CO2 into the atmosphere, which will act as a positive feedback, increasing CC. As a result, reducing atmospheric global warming gas emissions and improving SOC is a global problem. According to Brouder and Volenec [18], CC factors including temperature, precipitation (quantityand distribution), and atmospheric CO2 levels are expected to impact agricultural production patterns over the world. Changing precipitation patterns and greater global temperatures are projected to be connected with increased CO2 levels, and these variables may diminish or cancel any improvements in output or perhaps lower production below the critical levels. According to Bocchiola et al. [19], the effects of CC on agriculture may include (1) CO2 increasing in respiration, primarily for C3 plants, and (2) variations in temperature and rainfall, possibly leading to altered agricultural output over the twenty-first century.
However, according to Amouzou et al. [20], CC and variability threaten crops and food security in some areas. Production systems that are already vulnerable to soil fertility loss have become increasingly vulnerable to rainfall variability and climate change. These pressures will have the greatest effect on resource efficiency in cereal-based agricultural systems like maize (Zea mays) and sorghum (Sorghum bicolor). Brouder and Volenec [18] emphasized that any possible variations in agricultural productivity may result in related changes in the application of plant nutrients. Local possible productivity is influenced by dominant climate, surrounding CO2 levels, and crop features; however, this productivity is nearly continually controlled by the resources of the root zone (i.e., water and nutrients) and decreased by diseases and pests. They also emphasized that existing recommendations on nutrient management depend on the recognition of crop-specific requirements to achieve expected production and soil-specific nutrient provisions.
2.1. The Effect of CO2 Elevation
Elevated CO2 in the atmosphere has direct and indirect impacts on soil properties and processes via forced alterations in soil water and temperature, in addition to the competing of nutrients [9]. Investigations on plant response to the elevation of CO2 may not be representative of the conditions of actual fields in some plant species. There was a drawback in the effectiveness of the experiments of raised CO2 and temperature for determining their impacts on photosynthesis and other processes of the ecosystem. They attributed this to incorporating short-term sudden variations in elevated CO2 or temperature in the laboratory or field experiments. These changes rarely generated quantifiable variations in net primary productivity (NPP), C in the ecosystem, or other properties linked to long-standing responses to continuing CC [16]. Hatfield and Prueger [16] studied the influence of elevated CO2 on native grass and cultivated ecosystems and found that understanding the link between CC and plant nutrition may be best accomplished firstly by understanding this interaction in both range and grazing areas. Nitrogen is a limiting element for plant development in certain ecosystems. Terrestrial N exists in organic forms that are unavailable to plants and so the concept of how the response of rangeland to global CC will differ depending on the N cycling rate among the organic and inorganic N molecules being tested. Plant materials or roots that fall to the soil surface are degraded by fauna and microflora in the soil and so become a component of the soil organic matter (SOM) pool. Plant-available N forms and other minerals will be produced during the SOM decomposition stage. Many environmental and plant factors influence the rate of N-release, which is then impacted by CC and CO2 enrichment.
The interplay between CO2 and N in cultivated agroecosystems is as complex as it is in native ecosystems. The critical concentration of N for sufficient growth declined in several species when CO2 concentration increased. Hatfield and Prueger [16] supposed that one of the essential questions in this context was whether elevating CO2 concentrations influenced the plant nutrient concentrations. It is proposed that the required amount of N fertilizer in C3 crops for achieving the greatest productivity under elevated CO2 concentration would not increase because the critical concentration of N declines in C3 species with elevating CO2. On the other hand, in C4 plants, the required N for achieving maximum production could increase owing to no indication of N decreasing with elevating CO2. Although they recommended that not all these changes could be specifically attributed to elevating CO2, CO2 rising might have been a factor associated with this decline.
Nitrogen is an essential nutrient that influences plant development and physiology. It is also a key component of amino acids, protein, nucleic acids, and chlorophyll, helping to regulate C absorption and metabolism. It is indicated that elevated CO2 reduces leaf N content, in general, due to the diluting effect. Moreover, the decreased transpiration rate by elevated CO2 might cause a decrease in mass flow, reducing N absorption with the mass flow. Furthermore, the N concentration in plants may be connected to endogenous abscisic acid concentrations, which are influenced by variations in root development. Furthermore, low leaf N might enhance cytokinin levels under elevated CO2, encouraging stomatal opening. It is noteworthy that N can impact the accumulation of endogenous abscisic acid and the responses of the stomata to endogenous abscisic acid, thereby influencing the stomatal response to the stress of drought under elevated O2 growth conditions [21].
Seneweera and Norton [22] stated that the response of legumes to elevating CO2 varies between plant species and may reveal variable degrees of determinacy or the effects of nutrient limitations other than those chosen because of simulated growth. They added that P and Mo have been involved in limiting elevated CO2 responses in specific situations, though this might be counted as a typical drawback that may become worse if C and N supplies increase. Generally, elevated CO2 in the atmosphere induced fluctuations in temperature and precipitation, which lead to alterations in the vital soil processes that expose the soils to many kinds of degradation—physio-chemical degradation, salinization, erosion, reduced available water with changes in the soil, declined soil nutrient storage, and depletion of soil biodiversity. Such unfavorable changes pose a big threat to soil productivity, water and soil quality, and production system sustainability [17].
It was reported that increased CO2 reduced the levels of other nutrients in edible sections of crops, which were lower than N concentrations, such as Ca, K, Mg, and P. High CO2 levels have also been demonstrated to reduce plant concentrations of Co, Fe, Mg, Mn, Ni, S, and Zn [14][23][24]. However, Wang et al. [25] reported that although increased CO2 has a beneficial effect on agricultural productivity, warming has the capacity to negatively affect crop yield by reducing photosynthetic C absorption rates and shortening the growth period of plants. They added that it is unknown how variations in plant growth caused by CO2 elevation and/or warming impact will make soil micro-nutrients available and accessible to plants (i.e., it is not only controlled by its soil concentration but also by climatic conditions). However, Wang et al. [25] mentioned that elevated CO2 and warming together increased the availability of several soil micronutrients like Cu, Fe, and Zn. In addition, these climatic variables had a considerable effect on wheat’s micronutrient uptake and translocation. However, an increase in the availability of soil micronutrients did not necessarily increase micronutrient uptake (which is affected by the element types and crop growth stage).
2.2. The Effect of Temperature
High temperatures result in an increase in CO2 efflux from the soil. Global warming is lowering the SOC stock by boosting the rate of decomposition and, as a result, the residence time of C in the soil. As a result, soils are expected to be the main source of CO2 emissions in the future [26]. The increasing temperature modifies the factor of plant nutrients and different nutrient cycles in nature [9]. The rising temperature also influences physiological processes such as photosynthesis and respiration. The net impact of CC will vary owing to specific conditions. Therefore, in areas with cold spring and summer periods, where the growing season is short, higher temperatures may improve the crop yields. However, a warmer climate will reduce yields when the temperature increases. Additionally, increasing the temperature can shorten the growth and grain filling periods of the crop. Furthermore, various crop cultivars may display diverse responses to upcoming CC because of different seasons’ lengths [27]. Arndal et al. [13] stated that higher temperatures are likely to lengthen the growing season and enhance net mineralization, resulting in increased nutrient availability and perhaps increased uptake. Temperature increases caused by CC, which are both expected and observed to happen at faster rates at higher elevations, are believed to promote soil microbial activity and influence its physiology, resulting in higher levels of N mineralization and decomposing SOM at a quicker pace. This might result in releasing soil nutrients and their availability and decreased nutrient restriction. This happens more at warmer temperatures than colder ones [11][14].
The expected increases in temperature in the 21st century will likely influence the availability of nutrients in terrestrial ecosystems because biological pathways such as SOM decomposition and mineralization and N nitrification generally rise with increasing temperature [28][29]. Soils will be affected by CC through increasing the nutrient leaching rate and soil erosion [1], thus increasing nutrient depletion. Lotze-Campe [1] explained that higher temperatures affect nutrient conservation because warmer temperatures are possibly increasing the natural decomposition of SOM as a result of microbial activity stimulation. When mineralization is higher than plant uptake, the consequence will be the leaching of nutrients (Figure 2). It largely happens if plant requirements are low and nitrogen mineralization rates increase owing to rising soil temperatures. Wrage et al. [30] stated that P is an essential element for plant energy and sugar metabolism. Soil P availability is influenced by global CC. The P mineralization in litter tends to rise as temperatures rise. Temperature rises of 0.3 to 6.0 increased interest in mineralization by an average of 48%. Phosphatase exudation and plant P uptake are stimulated by higher levels of N. This might lead to higher soil P availability, which would be boosted further by enhanced P mobilization as a result of human activity. This will limit phytodiversity while promoting the spread of ruderal, fast growing plants [30].
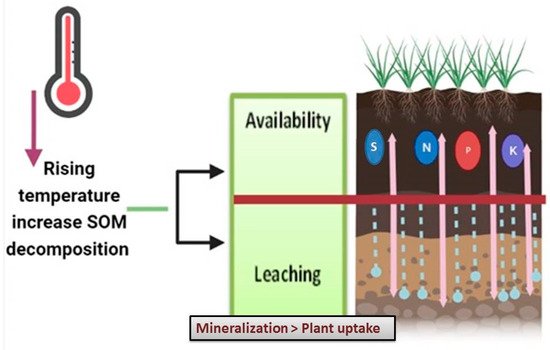
Figure 2. The effect of CC and rising temperature on SOM decomposition and thus soil nutrients.
Hatfield and Prueger [16] also reported that increasing temperatures accelerate the decomposition of SOM with much greater impacts in colder areas where minor elevations in temperature may have sensational influences on the decomposition of SOM plus other soil processes. On the other hand, increasing warming will interplay with soil water and increase its evaporation, which may constrain microbial activity. When the soil water becomes limiting, the interplay between temperature and soil water will grow and decomposition will be more reliant on the soil water content and less dependent on temperature. The low soil water content will decline the decomposition rates. Kirschbaum [31] reported that the temperature dependency of organic matter decomposition is significant in terms of ecophysiology, especially in light of potential CC feedback effects. It largely regulates whether or how much C will be emitted as a result of global warming, as well as the amount to which that C release constitutes a harmful positive feedback effect that results in more warming. Howden and Crimp [32] stated that climate is the main driver of cropping activities through significant impacts in the crop cycle involving soil moisture, droughts during the grain-filling period, heat damage, frost, and storm damages. Moreover, climate affects nutrient leaching, salt mobilization, and erosion risk. The CC may modify many of these impacts; for instance, the increasing temperature will lead to damage of grain numbers and quality, promoting drought stress and slashing initial soil water conditions. On the contrary, CC may also result in a decline in other hazards like dryland salinization.
2.3. The Effect of Precipitation
Global CC has already resulted in considerable variations in the volume, intensity, duration, and geography of rainfall. In addition, a large increase in the surface air temperature and seasonal patterns with similar or stronger moves are projected in the future [33]. Although rising temperatures are certain in the future, the projection of precipitation trends is still uncertain [29]. Precipitation changes and air temperature increases will substantially affect the prevailing temperature and moisture schemes of the root zone. The type and degree of the variations in both factors’ characteristics involve leaf area index and ground litter stores [18].
The considerable variations in precipitation directly influences the yield, especially in semi-arid and arid climates. Soil water is an essential resource in agriculture. Nutrient absorption is connected to soil water availability, which may be affected by climate change and changing precipitation patterns [34]. Additionally, nutrient leaching will also be forced by increased precipitation. It is known that intensive rainfall increases soil erosion, which is likely to be higher under CC. An increase of 1% in precipitation is projected to result in a 1.5 to 2% increase in erosion. Intensive rain and transforming from snowing to raining will moreover increase the erosion rate. Plant cover reduces soil erosion rates by weakening rain power (in addition to stabilizing soils by root and reducing sediment transportation by crop residue). Thus, variations in plant biomass can increase those impacts of soil erosion and nutrient leaching. On the other hand, in arid and semiarid climates, dry soil is susceptible to soil erosion across wind and rain [1]. Howden and Crimp [1] stated that commonly the integrating impacts of expected higher temperatures and decreased in-crop rainfall will lead to lower yields, an increase in the variability of yields, erosion risk, and thus the need for enhanced nutrient and water management. They also added that adaptation options in this regard include several different (separate or combined) levels of response, such as changing inputs (i.e., crop diversities and nutrient management) for matching the dominant climate, altering the amount and improving the proficiency of irrigation water, and more intensive residue and canopy management.
Climatic extremes in precipitation (such as floods and droughts) also cause damage to crop farming, which results in a reduction in the overall farm yields [1][2]. Rosenblatt and Schmitz [35] mentioned that although the precipitation is not projected to vary regularly over the world as CC continues, a consistent tendency over lots of terrestrial ecosystems is increasing the frequency of droughts, which has already doubled since 1970. Low water potential remains a widespread limiting factor of soil microbial activity (and thus nutrient mineralization) and in extremely dry soils may result in total inhibition in microbial activity. For example, the drought serious effect on mineralization is very distinct at high soil temperatures compared with low ones. Soil microorganisms physiologically respond to drought in many ways, such as a decline in cell osmotic potential, then by dehydration, and ultimately the death of microorganisms. In addition, Arndal et al. [13] reported that fine roots may dehydrate throughout a drought, reducing nutrient uptake and leading to declining root activity and nutrient mobility.
Generally, the fall of microbial activity in drought circumstances looks to be related to the drying length [29]. Rosenblatt and Schmitz [35] added that carbohydrate and protein levels generally increase in drought-stressed plants when osmotic potential decreases, allowing plants to retain more water. Furthermore, when drought stress increases, the relative water content of the leaves declines. However, the impacts of drought on plant nutrition are complex and vary depending on species, development stage, tissue type, drought severity, and length. Table 1 presents some studies that address the influence of climatic conditions on plant nutrition.
Table 1. Effect of climate variables on plant availability and uptake of plant nutrients.
| Climate Variable | Study Objective | The Effect on Plant | The Ecosystem | Ref. |
|---|---|---|---|---|
|
|
|
Two plants community predominated by Cassiope tetragona (L.) |
[36] |
|
|
|
Mediterranean forest, shrubland, and open areas | [12] |
|
Study the root nutrient uptake under predicted climate change |
|
Dry heathland/grassland | [13] |
|
Investigate the effect of CO2 enrichment and warming on the availability and plant uptake of soil micronutrients |
|
|
[25] |
|
Investigate the effect of environmental conditions on nutrient availability |
|
|
[11] |
|
Studying the effect of CO2 enrichment and canopy warming on nutrient concentration and accumulation (among other aims) |
|
|
[37] |
|
Examining the effect of climatic variables and pathogens on C, N, and P |
|
Mediterranean forest soil | [38] |
|
|
[39] | ||
|
|
|
Desert soil | [40] |
|
|
|
Eucalyptus globulus plants | [41] |
This entry is adapted from the peer-reviewed paper 10.3390/su14020914
References
- Lotze-Campe, H. Climate Change, Population Growth, and Crop Production: An Overview. In Crop Adaptation to Climate Change, 1st ed.; Yadav, S.S., Redden, R.J., Hatfield, J.L., Lotze-Campen, H., Hall, A.E., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2011.
- Hossain, M.S.; Arshad, M.; Qian, L.; Zhao, M.; Mehmood, Y.; Kächele, H. Economic impact of climate change on crop farming in Bangladesh: An application of Ricardian method. Ecol. Econ. 2019, 164, 106354.
- Elbasiouny, H.; Elbanna, B.A.; Al-Najoli, E.; Alsherief, A.; Negm, S.; El-Nour, E.A.; Nofal, A.; Sharabash, S. Agricultural Waste Management for Climate Change Mitigation: Some Implications to Egypt. In Waste Management in MENA Regions, Springer Water Book Series; Negm, A.M., Shareef, N., Eds.; Springer: Cham, Switzerland, 2020.
- Elbehiry, F.; Mahmoud, M.A.; Negm, A. Land Use in Egypt’s Coastal Lakes: Opportunities and Challenges. In Egyptian Coastal Lakes and Wetlands: Part I—Characteristics and Hydrodynamics; Negm, A.M., Bek, M.A., Abdel-Fattah, S., Eds.; Springer International Publishing AG: Cham, Switzerland, 2018.
- Shukla, S.K.; Shee, S.; Maity, S.K.; Solomon, S.; Awasthi, S.K.; Gaur, A.; Pathak, A.D.; Jaiswal, V.P. Soil Carbon Sequestration and Crop Yields in Rice–Wheat and Sugarcane–Ratoon–Wheat Cropping Systems Through Crop Residue Management and Inoculation of Trichoderma viride in Subtropical India. Sugar Tech 2016, 19, 347–358.
- Egli, M.; Bösiger, M.; Lamorski, K.; Sławiński, C.; Plötze, M.; Wiesenberg, G.L.; Tikhomirov, D.; Musso, A.; Hsu, S.-Y.; Raimondi, S. Pedogenesis and carbon sequestration in transformed agricultural soils of Sicily. Geoderma 2021, 402, 115355.
- Elbasiouny, H.; Elbehiry, F. Soil Carbon Sequestration for Climate Change Mitigation: Some Implications to Egypt. In Climate Change Impacts on Agriculture and Food Security in Egypt; Springer: Cham, Switzerland, 2020; pp. 151–181.
- Soares, J.C.; Santos, C.S.; Carvalho, S.M.P.; Pintado, M.M.; Vasconcelos, M.W. Preserving the nutritional quality of crop plants under a changing climate: Importance and strategies. Plant Soil 2019, 443, 1–26.
- Dotaniya, M.L.; Rajendiran, S.; Meena, B.P.; Meena, A.L.; Dotaniya, C.K.; Jat, R.L.; Saha, J.K. Elevated Carbon Dioxide (CO2) and Temperature vis-a-vis Carbon Sequestration Potential of Global Terrestrial Ecosystem. In Conservation Agriculture; Bisht, J.K., Ed.; Springer: Singapore, 2016.
- Gong, H.; Gao, J. Soil and climatic drivers of plant SLA (specific leaf area). Glob. Ecol. Conserv. 2019, 20, e00696.
- Koller, E.K.; Phoenix, G.K. Seasonal dynamics of soil and plant nutrients at three environmentally contrasting sites along a sub-Arctic catchment sequence. Polar Biol. 2017, 40, 1821–1834.
- Matias, L.; Castro, J.; Zamora, R. Soil-nutrient availability under a global-change scenario in a Mediterranean mountain ecosystem. Glob. Chang. Biol. 2010, 17, 1646–1657.
- Arndal, M.F.; Merrild, M.P.; Michelsen, A.; Schmidt, I.K.; Mikkelsen, T.N.; Beier, C. Net root growth and nutrient acquisition in response to predicted climate change in two contrasting heathland species. Plant Soil 2013, 369, 615–629.
- Pilbeam, D.J. Breeding crops for improved mineral nutrition under climate change conditions. J. Exp. Bot. 2015, 66, 3511–3521.
- Alshaal, T.; El-Ramady, H.; Al-Saeedi, A.H.; Shalaby, T.; Elsakhawy, T.; Omara, A.E.-D.; Gad, A.; Hamad, E.; El-Ghamry, A.; Mosa, A.; et al. The Rhizosphere and Plant Nutrition Under Climate Change. In Essential Plant Nutrients; Naeem, M., Ed.; Springer: Cham, Switzerland, 2017.
- Hatfield, J.L.; Prueger, J.H. Agroecology: Implications for Plant Response to Climate Change. In Crop Adaptation to Climate Change, 1st ed.; Yadav, S.S., Redden, R.J., Hatfield, J.L., Lotze-Campen, H., Hall, A.E., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2011.
- Sarfraz, R.; Hussain, A.; Sabir, A.; Fekih, I.B.; Ditta, A.; Xing, S. Role of biochar and plant growth promoting rhizobacteria to enhance soil carbon sequestration—A review. Environ. Monit. Assess. 2019, 191, 251.
- Brouder, S.M.; Volenec, J.J. Impact of climate change on crop nutrient and water use efficiencies. Physiol. Plant. 2008, 133, 705–724.
- Bocchiola, D.; Brunetti, L.; Soncini, A.; Polinelli, F.; Gianinetto, M. Impact of climate change on agricultural productivity and food security in the Himalayas: A case study in Nepal. Agric. Syst. 2019, 171, 113–125.
- Amouzou, K.A.; Lamers, J.P.; Naab, J.B.; Borgemeister, C.; Vlek, P.L.; Becker, M. Climate change impact on water- and nitrogen-use efficiencies and yields of maize and sorghum in the northern Benin dry savanna, West Africa. Field Crop. Res. 2019, 235, 104–117.
- Rajput, V.D.; Chen, Y.; Ayup, M. Effects of high salinity on physiological and anatomical indices in the early stages of Populus euphratica growth. Russ. J. Plant Physiol. 2015, 62, 229–236.
- Seneweera, S.; Norton, R.M. Plant Responses to Increased Carbon Dioxide. In Crop Adaptation to Climate Change, 1st ed.; Yadav, S.S., Redden, R.J., Hatfield, J.L., Lotze-Campen, H., Hall, A.E., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2011.
- Taub, D.R.; Wang, X. Why are nitrogen concentrations in plant tissues lower under elevated CO2? A critical examination of the hypotheses. J. Integr. Plant Biol. 2008, 50, 1365–1374.
- McGrath, J.M.; Lobell, D.B. Reduction of transpiration and altered nutrient allocation contribute to nutrient decline of crops grown in elevated CO2 concentrations. Plant Cell Environ. 2013, 36, 697–705.
- Wang, J.; Zhang, X.; Li, L.; Cheng, K.; Zheng, J.; Zheng, J.; Shen, M.; Liu, X.; Pan, G. Changes in micronutrient availability and plant uptake under simulated climate change in winter wheat field. J. Soils Sediments 2016, 16, 2666–2675.
- Shahzad, K.; Sintim, H.; Ahmad, F.; Abid, M.; Nasim, W. Importance of Carbon Sequestration in the Context of Climate Change. In Building Climate Resilience in Agriculture; Jatoi, W.N., Mubeen, M., Ahmad, A., Cheema, M.A., Lin, Z., Hashmi, M.Z., Eds.; Springer: Cham, Switzerland, 2022.
- Adavi, Z.; Moradi, R.; Saeidnejad, A.H.; Tadayon, M.R.; Mansouri, H. Assessment of potato response to climate change and adaptation strategies. Sci. Hortic. 2018, 228, 91–102.
- Cookson, W.; Osman, M.; Marschner, P.; Abaye, D.; Clark, I.; Murphy, D.; Stockdale, E.; Watson, C. Controls on soil nitrogen cycling and microbial community composition across land use and incubation temperature. Soil Biol. Biochem. 2007, 39, 744–756.
- Rennenberg, H.; Dannenmann, M.; Gessler, A.; Kreuzwieser, J.; Simon, J.; Papen, H. Nitrogen balance in forest soils: Nutritional limitation of plants under climate change stresses. Plant Biol. 2009, 11 (Suppl. 1), 4–23.
- Wrage, N.; Chapuis-Lardy, L.; Isselstein, J. Phosphorus, Plant Biodiversity and Climate Change. In Sociology, Organic Farming, Climate Change and Soil Science; Sustainable Agriculture Reviews 3; Springer: Dordrecht, The Netherlands, 2009.
- Kirschbaum, M.U.F. The temperature dependence of organic-matter decomposition—Still a topic of debate. Soil Biol. Biochem. 2006, 38, 2510–2518.
- Howden, S.M.; Crimp, S. Regional impacts: Australia. In Crop Adaptation to Climate Change; Yadav, S.S., Redden, R.J., Hatfield, J.L., Lotze-Campen, H., Hall, A.E., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2011; pp. 143–155.
- Simon, J.; Dannenmann, M.; Pena, R.; Gessler, A.; Rennenberg, H. Nitrogen nutrition of beech forests in a changing climate: Importance of plant-soil-microbe water, carbon, and nitrogen interactions. Plant Soil 2017, 418, 89–114.
- Khan, S.; Anwar, S.; Sun, M.; Shafiq, F.; Khalilzadeh, R.; Gao, Z.-Q. Characterizing Differences in Soil Water Content and Wheat Yield in Response to Tillage and Precipitation in the Dry, Normal, and Wet Years at the Loess Plateau. Int. J. Plant Prod. 2021, 15, 655–668.
- Rosenblatt, A.E.; Schmitz, O.J. Climate Change, Nutrition, and Bottom-Up and Top-Down Food Web Processes. Trends Ecol. Evol. 2016, 31, 965–975.
- Jonasson, S.; Michelsen, A.; Schmidt, I.K.; Nielsen, E.V. Responses in microbes and plants to changed temperature, nutrient, and light regimes in the arctic. Ecology 1999, 80, 1828–1843.
- Wang, J.; Li, L.; Lam, S.K.; Zhang, X.; Liu, X.; Pan, G. Changes in nutrient uptake and utilization by rice under simulated climate change conditions: A 2-year experiment in a paddy field. Agric. For. Meteorol. 2018, 250, 202–208.
- Avila, J.M.; Gallardo, A.; Gómez-Aparicio, L. Pathogen-induced tree mortality interacts with predicted climate change to alter soil respiration and nutrient availability in Mediterranean systems. Biogeochemistry 2019, 142, 53–71.
- Yang, X.; Zhang, P.; Wei, Z.; Liu, J.; Hu, X.; Liu, F. Effects of elevated CO2 and nitrogen supply on leaf gas exchange, plant water relations and nutrient uptake of tomato plants exposed to progressive soil drying. Sci. Hortic. 2021, 292, 110643.
- Luo, Y.; Peng, Q.; Li, K.; Gong, Y.; Liu, Y.; Han, W. Patterns of nitrogen and phosphorus stoichiometry among leaf, stem and root of desert plants and responses to climate and soil factors in Xinjiang, China. Catena 2020, 199, 105100.
- Macdonald, C.A.; Anderson, I.C.; Khachane, A.; Singh, B.P.; Barton, C.V.; Duursma, R.A.; Ellsworth, D.S.; Singh, B.K. Plant productivity is a key driver of soil respiration response to climate change in a nutrient-limited soil. Basic Appl. Ecol. 2020, 50, 155–168.
This entry is offline, you can click here to edit this entry!