Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Subjects:
Agriculture, Dairy & Animal Science
Polycomb repressive complex 2 (PRC2) represents a group of evolutionarily conserved multi-subunit complexes that repress gene transcription by introducing trimethylation of lysine 27 on histone 3 (H3K27me3). PRC2 activity is of key importance for cell identity specification and developmental phase transitions in animals and plants.
- polycomb
- PRC2
- H3K27me3
- evolution
- green lineage
- plant
1. Introduction
DNA in the eukaryotic nucleus winds around octamers of histones, forming nucleosomes, the basic units of chromatin. DNA and histones are subject to chemical modifications, such as methylation, phosphorylation, acetylation, and others, which are instructive for or correlate with chromatin structure. Remodeling of chromatin structure between more open (accessible) or more compact (inaccessible) states by chromatin-modifying and -remodeling complexes governs the distribution of DNA in the nuclear space and allows for gene activation or repression, respectively.
Among crucial modulators of chromatin structure are polycomb group (PcG) proteins, which form multi-subunit polycomb repressive complexes (PRCs) (reviewed in the work of [1,2,3,4,5]). Major PRCs are histone-modifying complexes that confer different and even counteracting enzymatic activities, which mediate gene repression. In animals and plants, PRC1 is an E3 ubiquitin ligase complex that catalyzes histone 2A lysine ubiquitination (H2AKub: K118 in Drosophila, K119 in vertebrates, and K121 in plants) [6,7,8], and PRC2 is a histone methyltransferase (HMT) complex that catalyzes histone 3 lysine 27 methylation (H3K27me) [9]. Some PRCs catalyze H2Aub removal. In animals, the PR-DUB (polycomb repressive deubiquitinase) complex catalyzes histone H2A deubiquitination, which contributes to gene repression (reviewed in the work of [10,11]). Similarly, in plants, H2A deubiquitinases UBP12 and UBP13 are implicated in polycomb repression [12,13], but the composition of associated protein complexes remains enigmatic. Additional PRCs have been described in animals, such as Drosophila pleiohomeotic repressive complex (PhoRC) that does not confer enzymatic activity ([14], reviewed in the work of [15]). As histone modifications introduced by PRCs are heritable during mitotic cell divisions (reviewed in the work of [16,17,18]), PRCs provide an epigenetic memory system required for stable cell identity, for an adequate response to external cues, and even for stable repression of genomic repeats (reviewed in the work of [19]). In line with its function in cell identity maintenance in animals, PRC2 dysfunction is frequently associated with cancer development and PRC2 is a potent target for anticancer therapy (reviewed in the work of [20,21,22]).
PRC1 and PRC2 are conserved in animal and plant models. PRC1 was long considered to be animal specific since a homolog of Polycomb (Pc), the Drosophila PRC1-defining protein subunit [23], is missing in plants. Later, orthologs of PRC1 catalytic subunits RING1 (Really Interesting New Gene 1) and BMI1 (B cell-specific Moloney murine leukemia virus integration site 1) were identified in plants and worms [24]. RING1 and BMI1 were shown to be indispensable for plant development and required for H2Aub [7,25,26]. RING1 and BMI1 orthologs, as well as plant-specific PRC1 subunits, have been found in different plants species ([27], reviewed in the work of [28]). Hence, PRC1 is present in plants, although its core composition differs from animals and may have originated through convergent evolution (reviewed in the work of [28,29,30]). In contrast to PRC1, PRC2 core composition and H3K27me activity are well conserved throughout eukaryotic lineages, and PRC2 is hypothesized to have emerged through divergent evolution [31]. The evolution of PRC2 has been shaped by genome duplication and subfunctionalization, and the number of different PRC2 complexes tends to rise with increasing body plan complexity (Figure 1) ([32], reviewed in the work of [15,33,34]). In animals, PRC2 catalyzes H3K27 mono, di, and trimethylation (i.e., H3K27me1, H3K27me2, H3K27me3), but in flowering plants, it catalyzes H3K27me3 ([35,36], reviewed in the work of [9,18]). In model species of both animals and plants, H3K27me3 is largely associated with transcriptional silencing of developmental genes ([37,38], reviewed in the work of [39,40]). PRC2 composition, its biochemical and developmental functions are well studied in animal and in flowering plant model species, and we refer to recent reviews for detailed information [41,42,43,44,45,46,47,48,49].
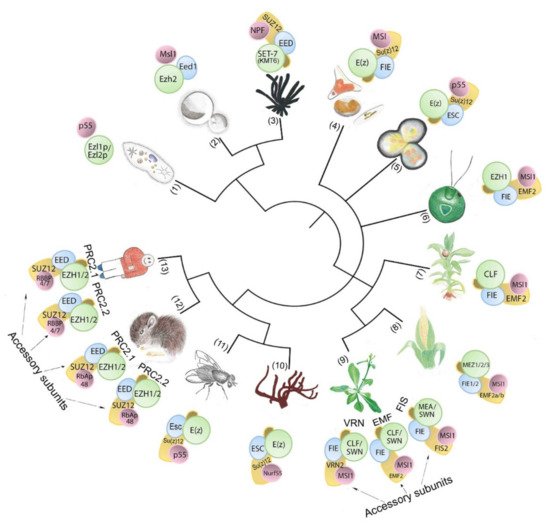
Figure 1. PRC2-core subunit diversity in unicellular and multicellular organisms. The number of subunit homologs and putative PRC2 complexity hypothetically increases with body plan complexity. Numbers in brackets indicate species (1) Paramecium tetraurelia; (2) Cryptococcus neoformans; (3) Neurospora crassa; (4) Phaeodactylum tricornutum; (5) Cyanidioschyzon merolae; (6) Chlamydomonas reinhardtii; (7) Physcomitrium patens; (8) Zea mays; (9) Arabidopsis thaliana; (10) Amphimedon queenslandica; (11) Drosophila melanogaster; (12) Mus musculus; (13) Homo sapiens..
2. Features of PRC2 Core Composition and Function Are Conserved in Animal and Plant Models
The core of PRC2 is generally composed of four protein subunits that are conserved in multicellular eukaryotic model organisms (Figure 1, Table 1). The four subunits enhancer of zeste (E(z)), suppressor of zeste 12 (Su(z)12), extra sex combs (Esc), and nucleosome remodeling factor (Nurf55, also called p55) are the essential PRC2 components present in D. melanogaster (Figure 2) ([50], reviewed in the work of [15]). E(z) contains the catalytic SET domain (Suppressor of variegation 3–9 (Su(var)3–9), Enhancer of zeste (E(z)), Trithorax (Trx) domain), which is responsible for the HMT activity ([50], reviewed in the work of [51]). Su(z)12 is a VEFS (VRN2-EMF2-FIS2-Su(z)12) domain-containing protein, and Esc and p55/Nurf55 are two WD40 repeat (WDR) domain proteins. Su(z)12, Esc, and Nurf55 are non-catalytic subunits that are crucial for PRC2 catalytic activity ([52,53,54,55], reviewed in the work of [56]). PRC2 containing all four core components is 1000 times more active than the E(z) subunit alone [50,57]. Esc and Su(z)12 play critical roles in stimulating the HMTase activity of E(z) [58,59], and the loss of Esc impairs global H3K27me in Drosophila embryos [52,57]. Homologs of Drosophila PRC2 subunits are conserved in mammals, fungi, plants, red algae, and diatoms (Figure 1, Table 1) ([60,61,62,63,64,65], reviewed in the work of [66,67,68,69]). In mammals, the homologs of E(z), Su(z)12, Esc, and p55 are EZH1 and EZH2, SUZ12, EMBRYONIC ECTODERM DEVELOPMENT (EED), and RETINOBLASTOMA BINDING PROTEINS (RBBP4 or RBBP7), respectively. Minimal PRC2 in mammals comprises the EZH1/2, EED, and SUZ12 subunits in a 1:1:1 stoichiometry [70], and the trimeric core is responsible for H3K27me1/2/3 (reviewed in the work of [71]). RBBP4 or RBBP7, which display 92% sequence identity, are frequently found together in the same complex, but they may differ functionally, as only RBBP4 is involved in maintaining stem cell identify ([72], reviewed in the work of [73]). Core PRC2 possesses lower activity and/or affinity to target sites unless associated with accessory proteins (reviewed in the work of [56]) that define subtypes of PRC2s ([74], reviewed in the work of [75,76]). In humans, two PRC2 subtypes are present based on association with accessory proteins, namely PRC2.1 (contains polycomb-like 1–3 (PCL1/2/3), elongin BC and PRC2 (EPOP), PRC2-associated LCOR isoform 1/2 (PALI1/2)), and PRC2.2 (contains jumonji and AT-rich interaction domain-containing 2 (JARID2), adipocyte enhancer-binding protein (AEBP2)) (reviewed in the work of [10]). The molecular structures have been determined for Drosophila, human, mouse, and the fungus Chaetomium thermophilum PRC2 (Figure 2) ([60,77,78,79,80], reviewed in the work of [34,81,82,83,84]).
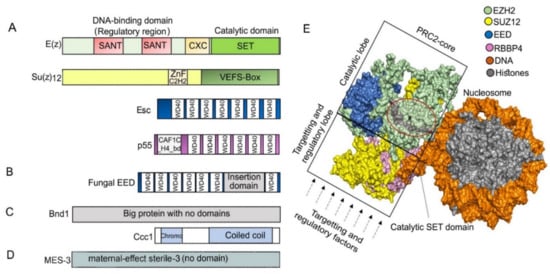
Figure 2. (A) Typical domain architecture of PRC2 core subunits in D. melanogaster. The catalytic subunit E(z) contains SANT (SWI3-ADA2-N-CoR-TFIIIB), CXC (pre-SET domain with C-X(6)-C-X(3)-C-X-C motif), and SET (Su(var)3–9, enhancer of zeste, and trithorax domain) domains, and it catalyzes H3K27me through the SET domain. Su(z)12 contains ZnF_C2H2 (cysteine 2-histidine 2 zinc finger domain) and VEFS (VRN2-EMF2-FIS2-Su(z)12) domain and is responsible for assembling the PRC2 complex, while Esc (WD40 repeat protein) helps to stabilize and enhance E(z) activity. P55 (WD40 repeat protein) can bind to histones and Su(z)12 and is necessary for nucleosome interaction. (B) Most of the studied fungal Esc homologs EED carry a long insertion domain in their C-terminal part. (C) While Su(z)12 is missing, subunits Bnd1 (Big protein with no domains) and Ccc1 (Chromodomain and a coiled coil region) are present in Cryptococcus neoformans. (D) Su(z)12 is missing in C. elegans, but the protein MES-3 (no detectable domains) is a part of the core PRC2. (E) PRC2 core structure. Surface depiction of human PRC2 core complex was created using PyMOL 2.4.1 [85] based on PDB ID: 6WKR [86]; only core subunits are shown. In a catalytically active complex, the EED subunit is encircled by EZH2, and the C-terminal VEFS domain of SUZ12 is sandwiched between the EED and EZH2-SET domain [87]. The region in the circle highlights the lysine binding channel (gray) through which a methyl group is transferred from the cofactor S-adenosylmethionine (SAM)) to the substrate H3 peptides. The catalytic or targeting and regulatory functions of mammalian PRC2 are separated into two regions within the structure: into the top catalytic lobe (containing EZH, VEFS domain of SUZ12 and EED) and the bottom targeting and regulatory lobe (containing RBBP4/7 and N-terminal SUZ12) (reviewed in the works of [42,46,84,88]).
Table 1. The distribution of PRC2 core components in different species. (Black ✓ indicates prediction and green ✓ indicates experimental evidence; numbers in brackets indicate the number of homologs identified, ‘?’ indicates that the presence or absence is still unclear, and ‘-’ indicates absence.)
| Supergroups | Eukaryotic Group/Kingdom | Phylum or Class | Species | E(z) | Su(z)12 | Esc | p55 | References |
|---|---|---|---|---|---|---|---|---|
| Archaeplastida | Eukaryota | Rhodophyta | Cyanidioschyzon merolae | ✓ | ✓ | ✓ | ✓ (2) | [63] |
| Viridiplantae | Chlorophyta | Ostreococcus lucimarinus | ✓ | ✓ | ✓ | ✓(2) | [89] | |
| Chlamydomonas reinhardtii | ✓ | ✓ | ✓(2) | ✓(2) | [63,89] | |||
| Volvox carteri | ✓(2) | ? | ✓ | ✓(2) | [89] | |||
| Viridiplantae—Embryophyta | Bryophyta-Bryopsida | Physcomitrium patens | ✓ | ✓(3) | ✓ | ✓(2) | [61,62,89,90] | |
| Lycophyte-Lycopodiopsida | Selaginella moellendorffii | ✓(2) | ✓ | ✓ | ✓(3) | [89,90,91] | ||
| Gymnosperm | Picea abies | ✓ | ? | ? | ? | [92] | ||
| Angiosperms-Monocot | Oryza sativa | ✓(2) | ✓(2) | ✓(2) | ✓(2) | [91,93,94,95,96] | ||
| Triticum aestivum | ✓(9) | ✓(8) | ✓ (7) | ✓(6) | [97] | |||
| Zea mays | ✓(3) | ✓(2) | ✓(2) | ✓(5) | [94] | |||
| Brachypodium distachyon | ✓(2) | ✓(2) | ✓(3) | ✓(4) | [89,98,99] | |||
| Hordeum vulgare | ✓(3) | ✓(3) | ✓ | ✓(2) | [94,100,101] | |||
| Sorghum bicolor | ✓(2) | ✓(3) | ✓(2) | ✓(2) | [94] | |||
| Angiosperms-Eudicot | Arabidopsis thaliana | ✓(3) | ✓(3) | ✓ | ✓(5) | [102] | ||
| Chromalveolata | SAR—Stramenopiles | Bacillariophyceae (diatoms) | Phaeodactylum tricornutum | ✓ | ✓ | ✓ | ✓ | [89,103] |
| Ochrophyta-Phaeophyceae | Ectocarpus | - | - | - | - | [104] | ||
| SAR—Alveolata | Cilliophora/cilliates | Paramecium tetraurelia | ✓(2) | - | - | ✓ | [31,105,106] | |
| Tetrahymena thermophila | ✓ | ✓ | ✓ | ✓ | [107,108] | |||
| Opisthokonta | Fungi | Basidiomycota | Cryptococcus neoformans | ✓ | - | ✓ | ✓ | [64,109] |
| Ascomycota | Fusarium graminearum | ✓ | ✓ | ✓ | ✓ | [109,110] | ||
| Chaetomium thermophilum | ✓ | ✓ | ✓ | ✓ | [79] | |||
| Neurospora crassa | ✓ | ✓ | ✓ | ✓ | [109,111,112] | |||
| Saccharomyces cerevisiae | - | - | - | - | [31,75] | |||
| Filasterea—single-celled eukaryote | Capsaspora | Capsaspora owczarzaki | - | - | - | - | [113] | |
| Animalia/animals | Porifera | Amphimedon queenslandica | ✓(4) | ✓ | ✓(2) | ✓ | [114] | |
| Cnidaria/Hydrozoa | Hydra vulgaris | ✓ | ✓ | ✓ | ✓ | [115,116,117,118] | ||
| Insecta/insects | Drosophila melanogaster | ✓ | ✓ | ✓ | ✓ | [119,120,121] | ||
| Nematoda/nematodes | Caenorhabditis elegans | ✓ | - | ✓ | - | [122,123] | ||
| Reptilia/reptiles | Anolis carolinensis | ✓ | ✓ | ✓ | ✓ | [124,125,126,127] | ||
| Mammalia/mammals | Homo sapiens | ✓(2) | ✓ | ✓ | ✓ | [56,60] | ||
| Mus musculus | ✓(2) | ✓ | ✓ | ✓ | [128,129] |
The flowering dicot model plant Arabidopsis thaliana has three E(z) paralogs (CURLY LEAF (CLF), SWINGER (SWN) and MEDEA (MEA)), three Su(z)12 paralogs (EMBRYONIC FLOWER 2 (EMF2), VERNALIZATION 2 (VRN2), and FERTILIZATION INDEPENDENT SEED 2 (FIS2)), one ESC homolog (FERTILIZATION INDEPENDENT ENDOSPERM (FIE)), and five p55 homologs (MULTICOPY SUPPRESSOR OF IRA (MSI1–MSI5)), of which only MSI1 is known to be present in PRC2 ([102,130,131], reviewed in the works of [29,132]). SWN and CLF play significant roles during vegetative development and phase transitions ([133,134,135,136,137,138], reviewed in the work of [47,48]), while MEA is required during gametophyte development and early embryogenesis [131,139]. Three PRC2 complexes have been identified in Arabidopsis defined by the presence of respective Su(z)12 homologs: EMBRYONIC FLOWER (EMF), VERNALIZATION (VRN), and FERTILIZATION INDEPENDENT SEED (FIS). The EMF and VRN complexes are associated with either CLF or SWN, and the FIS complex contains MEA or SWN as a catalytic subunit ([140,141,142], reviewed in the work of [3,69]). In A. thaliana, accessory subunits that can physically interact with core PRC2 subunits have been identified, including several transcription factors, such as the ASYMETRIC LEAVES (AS), TELOMERE REPEAT BINDING FACTORS (TRBs), VIVIPAROUS1/ABI3-LIKE (VAL), additional PRC-chromatin-associated enzymatic or scaffold proteins, such as LIKE HETEROCHROMATIN PROTEIN 1 (LHP1), VERNALIZATION INSENSITIVE 3 (VIN3), VRN5, PWWP-DOMAIN INTERACTOR OF POLYCOMBS 1 (PWO1), INCURVATA11 (ICU11), components of the E3 ubiquitin ligase complexes, components of the DNA replication and chromosome segregation machineries (CTF4) or domesticated transposases (ALP1, ALP2) ([143,144,145,146,147,148,149], reviewed in the work of [3,33,45]). In spite of the well-described impact of PRC2 function on plant development, plant PRC2 composition and potential subcomplex function remain enigmatic. Additionally, structural and detailed biochemical information on plant PRC2 is not yet available, limiting detailed insight into PRC2 subunit interaction and mechanism of action in plants.
In animals, PRC2 catalyzes H3K27me1/2/3 [50], while in plants, H3K27me1 in centromeres and pericentromeres is catalyzed by the ARABIDOPSIS TRITHORAX-RELATED HMTs (ATXR5 and ATXR6) [150]. H3K27me3 is, therefore, the conserved hallmark of PRC2 enzymatic activity (Figure 3). H3K27me3 in D. melanogaster occupies broad domains that typically span more than 10 kb [151]. In mammals, H3K27me3 has two distribution patterns: large domains (>100 kb) encompassing the Hox loci and smaller domains of a few kilobases (reviewed by the authors of [75]). In addition to genic loci, H3K27me3 and PRC2 reside in poised enhancers (PEs) that often associate with bivalent genes in vertebrate pluripotent cells [152]. While PRC1 contributes to the PE marking globally [152] and also targets active enhancers in cancer cells [153], PRC2 is involved at PEs at specific loci [152]. Unlike in animals, the distribution of H3K27me3 in A. thaliana is usually limited to single genes, with the modification covering proximal parts of promoters and gene bodies with average enrichment regions of 1–1.5 kb [37,38,154,155]. Gene-limited distribution is also found in metabolic gene clusters that are organized in an operon-like manner [156]. In A. thaliana, H3K27me3 decorates approximately 20–30% of all protein-coding and miRNA genes [37,38,157] that are associated with plant development, hormone metabolism and response, but also with nutrient homeostasis [158,159], stress response ([160,161], reviewed in the works of [5,162,163]) or primary and secondary metabolism [156,161]. The potential for targeting is nevertheless more extensive, as 64% of protein-coding genes were identified as H3K27me3 targets when combining different timepoints of A. thaliana shoot apical meristem development during the transition to flowering [164].
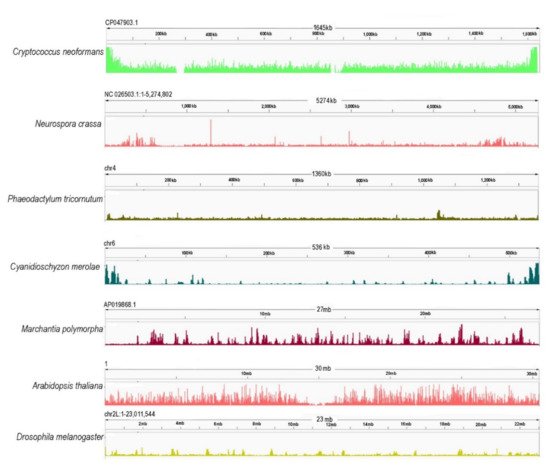
Figure 3. Chromosome-wide distribution of H3K27me3 in different model species [Cryptococcus neoformans (ASM1180120v1/PRJNA261445) [64], Neurospora crassa (NC12/PRJNA192863) [111], Phaedactylum tricornutum (ASM15095v2/PRJNA282957) [165], Cyanidioschyzon merolae (ASM9120v1/PRJNA362822) [63], Marchantia polymorpha (ASM993635v2/PRJNA553138) [166], Arabidopsis thaliana (TAIR10/PRJNA277409) [167], Drosophila melanogaster (PRJNA379297) [168]. Genome reference and accessions are given in brackets. Here, the publically available data were downloaded, cleaned by removing the library adapters, small reads (30), and low-quality (20) reads using Trim galore [169]. To map the sequenced data, Bowtie2 [170] was used with default parameters. Only mapped reads were kept with a quality threshold of 25. The filtering was performed using SAMTOOLS [171]. For the visualization of the data, IGV [172] was used, and for the parsing of the BAM files to BW files, BamCoverage—deeptools [173].
PRC2 itself lacks sequence-specific DNA-binding ability and therefore relies on accessory proteins for targeting specific loci. Several mechanisms by which PRC2 is recruited to the chromatin targets have been identified. Among these are transcription factor site-specific recruitment, interaction with RNAs, or association with chromatin features (reviewed in the work of [88]). In D. melanogaster, PRC2 is recruited to DNA targets by different transcription factors that interact with sequence motifs known as polycomb response elements (PREs) ([174,175], reviewed in the work of [176]). PREs in Drosophila are found in proximal promoter regions of developmental genes. They typically span about 1.5 kb [177] and contain numerous binding sites for a variety of DNA sequence-specific binding factors, such as the GTGT-motif-binding Combgap (Cg) [178], which together mediate PRC2 recruitment to target sites (reviewed in the work of [75]). Typical fly-like PREs are not found in mammals, and even though several mammalian PREs have been reported [179], they are not evolutionarily conserved. CpG islands, hypomethylated CG-rich chromatin regions of 1–2 kb, are associated with PRC2 recruitment in mammals, where accessory subunits rather than transcription factors mediate the recruitment ([180,181,182], reviewed in the work of [71]). Transcription factor-bound PRE-like elements also contribute to PRC2 recruitment in A. thaliana (reviewed in the work of [5]). Following the identification of PRC2-recruiting cis elements in the promoters of the KNOX genes [183] and the LEC2 gene [184], PRE-like elements (including GAGA motif, W-box, RY motif, GCCGCC motif, telobox motif, and others) bound by transcription factors were shown to contribute to PRC2 recruitment genome-wide [185]. Several transcription factors have been demonstrated to interact directly with PRC2 subunits to mediate the complex’s recruitment [144,146,183,186]. For instance, transcriptional repressors VIVIPAROUS1/ABI3-LIKE1 (VAL1) and VAL2 or TELOMERE REPEAT-BINDING FACTORS (TRBs) physically interact with SWN and CLF to recruit PRC2 to specific sequence motifs present in the promoters of target genes [144,146,187]. PRC2 recruitment through interaction with trans-acting factors that recognize cis elements is thus conserved in both plant as well as animal models (reviewed in the work of [45]).
Noncoding RNAs (ncRNAs), in particular long noncoding RNAs (lncRNAs), and nascent RNAs have also been implemented in PRC2 binding and recruitment to target sites (reviewed in the work of [188,189]). The formation of DNA-RNA hybrid structures (R-loops) is suggested to promote PRC2 recruitment in mammalian cells [190]. R-loops have been found at a variety of polycomb target gene loci, mostly found adjacent to the promoters (reviewed in the work of [191]), and interestingly, R-loop formation was detected at about one-third of PREs in Drosophila embryos [192], suggesting conservation of the mechanism. R-loops have been shown to positively and negatively impact recruitment, and their role is discussed [190,193]. lncRNAs are also known to contribute to PRC2 recruitment in A. thaliana. For instance, COLD ASSISTED INTRONIC NONCODING RNA (COLDAIR), COOLAIR, and COLDWRAP act in repressing FLOWERING LOCUS C (FLC) transcription by H3K27me3 [194,195] and the lncRNA APOLO contributes to LHP1 recruitment, H3K27me3 enrichment, and chromatin looping at the PINOID (PID) locus that encodes a key polar auxin transport regulator [196].
Last but not least, PRC2 recruitment and activity are prevented or promoted by its interaction with the existing chromatin environment (Figure 4). Several pre-existing chromatin modifications, including H3K27ac, H3K4me3, H3K36me2/3, and cytosine methylation in CpG islands, prevent PRC2 recruitment and/or inhibit its activity ([77,180,197,198,199,200], reviewed in the work of [201]). In addition to opposing PRC2, H3K36me3 may promote H3K27me3. In mouse embryonic stem cells, Phf19, a PCL ortholog, binds to H3K36me2/H3K36me3, recruiting PRC2 and lysine demethylases to promote PRC2 activity [202]. Although biochemical evidence is limited in plants [198], H3K27me3 mainly occupies regions that are depleted for active chromatin modifications [154,155]. An exception to this is regions of bivalent chromatin where activating and repressive modifications co-localize to potentiate rapid change of gene transcription. In both plant and animals, the best well-described bivalent chromatin is marked by H3K27me3 and H3K4me3 ([155], reviewed in the work of [203]), but other active modifications may co-localize with H3K27me3 including H3K4me1 in Brassica napus [204], or H3K18ac in the camalexin biosynthesis genes in A. thaliana [205]. In mammals, H3K27me3 is promoted in a self-reinforcement loop. Pre-existing H3K27me3 is bound by EED, which allosterically stimulates PRC2 to methylate adjacent unmodified H3K27 and promote the spreading of H3K27me3 [87,206,207]. H3K27me3 binding activity has not been shown for the A. thaliana EED homolog FIE. Nevertheless, A. thaliana MSI1 interacts with LHP1, which binds H3K27me3, and through this may enable H3K27me3 spreading and/or post-replicative maintenance of H3K27me3 [102,208]. In addition, H2AKub, the catalytic product of PRC1, can act as a recruitment platform for PRC2 ([209,210], reviewed in the works of [45,211]). The human PRC2 cofactors JARID2 and AEBP2 bind to H2AK119ub, triggering a positive feedback loop [77] that ensures the maintenance of transcriptional repression. In animals as well as in plants, genome-wide deposition of H3K27me3 and H2Aub seem to be partially dependent on each other. In animal models (Drosophila, mouse, and human cells), most studies suggest that H2Aub modification is independent of H3K27me3 deposition, while H3K27me3 levels are decreased upon disruption of H2Aub (reviewed in the work of [212]). In A. thaliana, H3K27me3 overlaps with H2AK121ub at a subset of loci [26]. H2Aub is more widespread than H3K27me3, and its deposition is largely independent of PRC2 activity. In contrast, H3K27me3 depends on H2Aub at sites that carry both the marks, together indicating that PRC1 may be instructive for H3K27me3 in plants rather than vice versa [26]. Recently, three different chromatin states occupied either by H3K27me3 only, H2Aub only, or both modifications were described in A. thaliana, showing that accessibility increases from the inaccessible H3K27me3-only-marked chromatin to H3K27me3/H2Aub and H2Aub-only chromatin, that are mainly located at transcriptional hotspots [213]. This is in line with findings that H2Aub associates with responsive genes and its repressive function relies on H3K27me3 deposition and that H2A deubiquitination by UBP12 and UBP13 are required for stable H3K27me3-mediated repression [12,13]. Thus, while PRC1 and PRC2 activities both contribute to decreased chromatin accessibility, H3K27me3 seems to be a major contributor to stable gene repression within inaccessible chromatin regions in A. thaliana.
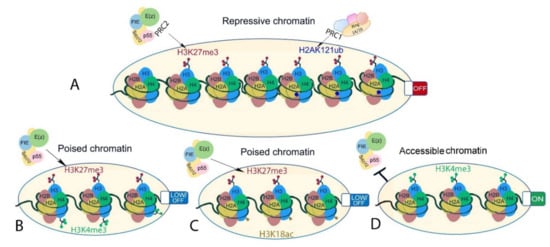
Figure 4. Different chromatin states associated or interacting with H3K27me3 in plants. (A) Facultative heterochromatin-inaccessible H3K27me3-marked chromatin (left) and inaccessible but permissive H3K27me3- and H2AK121ub-marked chromatin (right). (B,C) bivalent chromatin is marked by histone modifications with opposing effects on gene transcription. Examples are (B) H3K27me3-H3K4me3 and (C) H3K27me3 and H3K18ac. These are poised states where gene transcription is readily initiated upon stimuli. (D) Euchromatin showing chromatin state at transcriptionally active genes that is known to interfere with PRC2-mediated H3K27me3 deposition on the same histone tail in A. thaliana.
This entry is adapted from the peer-reviewed paper 10.3390/epigenomes6010003
This entry is offline, you can click here to edit this entry!