This entry focuses on recently characterized traits of the aquatic floating plant Lemna with an emphasis on its capacity to combine rapid growth with the accumulation of high levels of the essential human micronutrient zeaxanthin, due to an unusual pigment composition not seen in other fast-growing plants. In addition, Lemna’s response to elevated atmospheric carbon dioxide was evaluated in the context of the source–sink balance between plant sugar production and consumption. These and other traits of Lemnaceae are compared with those of other floating aquatic plants as well as terrestrial plants adapted to different environments. It was concluded that the unique features of aquatic plants reflect adaptations to the freshwater environment including rapid growth, high productivity, and exceptionally strong accumulation of high-quality vegetative storage protein and essential human antioxidant micronutrients. It was further concluded that the insensitivity of growth rate to environmental conditions and plant source–sink imbalance may allow duckweeds to take advantage of elevated atmospheric carbon dioxide levels via particularly strong stimulation of biomass production and either no (under favorable nutrient supply), or only minor (under limiting nutrient supply), declines in the growth of new tissue.
- chlorophyll fluorescence
- electron transport chain
- inflammation
- lutein
- photosystem
- photosynthetic capacity
- relative growth rate
1. Introduction
2. Exceptions to Common Trade-Offs: Araceae and Lemnaceae
2.1. Trade-Off between the Ability to Grow in Deep Shade and Full Sun
2.2. Common Trade-Off between Fast Growth and Antioxidant Accumulation
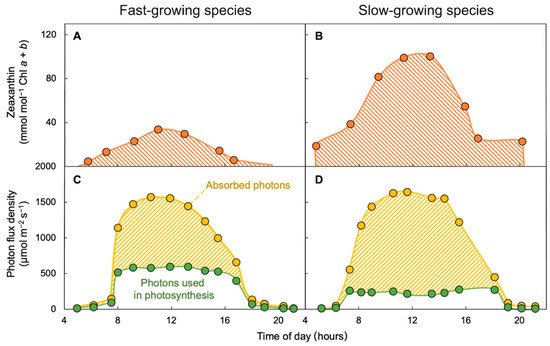
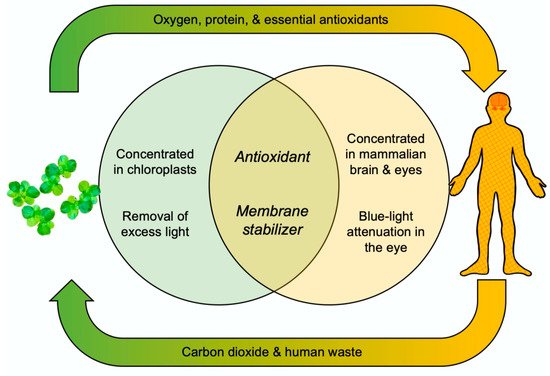
2.3. Zeaxanthin—Essential Human Micronutrient and Hard to Come by in the Diet
2.4. Lemna: An Unusual, Fast-Growing Hyperaccumulator of Zeaxanthin
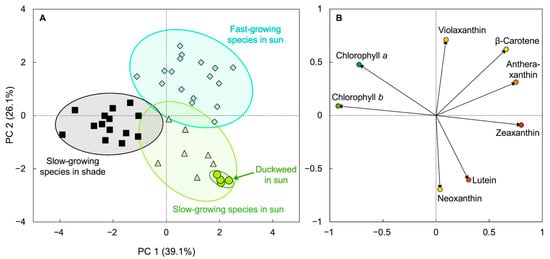
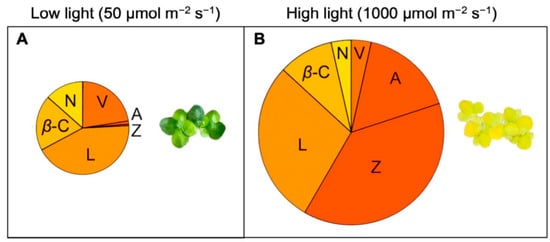
2.5. Remote Sensing of Duckweed Zeaxanthin Content and Biomass Production
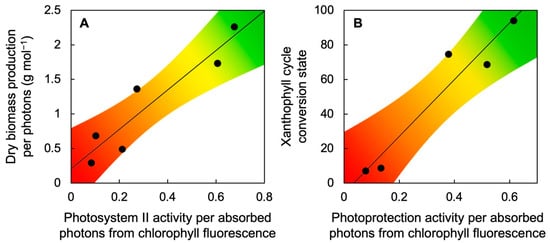
3. Comparative Evaluation of Adaptations in Aquatic and Terrestrial Plants
3.1. Rapid Growth in Duckweeds and Other Aquatic Plants
3.2. Continuum of Plant Adaptations to Water Availability
| Habitat | Water-Acquisition Strategy | Growth Pattern | Example |
|---|---|---|---|
| Aquatic plants | Continuous access to water body until it dries up | Fast growth | Lemnaceae |
| Terrestrial plants | Continuous access to water until source dries up | Very fast growth and life cycle completion | Desert ephemerals |
| Continuous access to water, increased root volume, osmotic adjustment | Relatively fast growth throughout life cycle | Annuals and biennials | |
| Continuous access to water table | Steady growth throughout the seasons | Palm and mesquite | |
| Enhanced acquisition of minimal soil water via large root volume, osmotic adjustments | Slow growth despite minimal soil water | Desert shrub Encelia, creosote bush |
|
| Storage of water in plants | Very slow growth | Succulents, cacti | |
| Tolerance of seasonal loss of water | Seasonal complete growth arrest | Conifers in frozen soil |
3.3. Plant Source–Sink Balance and the Response to Light, CO2, and Nutrient Supply
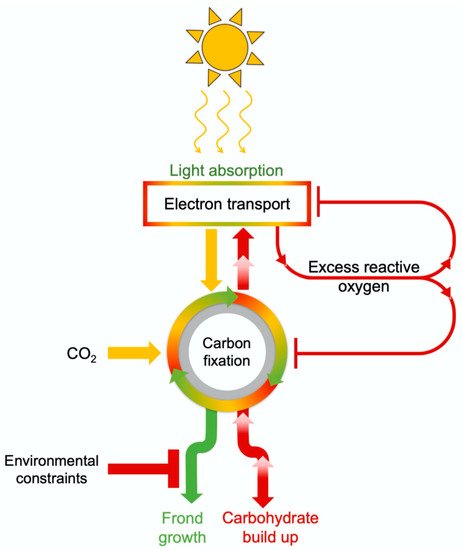
3.4. Comparative Evaluation of Plant Response to Light Supply
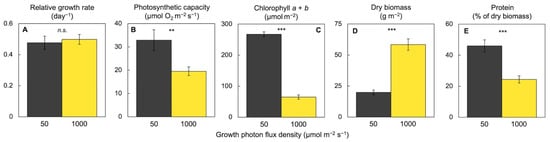
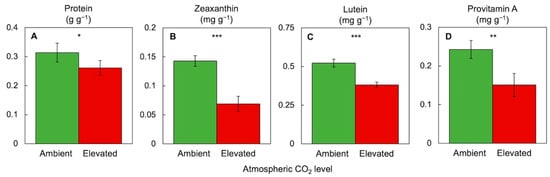
3.5. Comparative Evaluation of Plant Response to a Combination of High CO2, High Light Supply, and Limiting Mineral Nutrient Supply
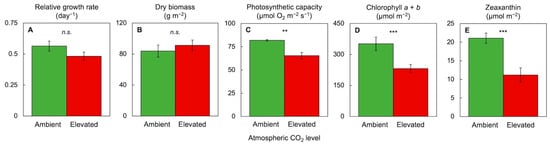
3.6. Comparative Evaluation of Plant Response to Combinations of High CO2, High Light Supply, and High Mineral Nutrient Supply
This entry is adapted from the peer-reviewed paper 10.3390/plants11020145
References
- Acosta, K.; Appenroth, K.J.; Borisjuk, L.; Edelman, M.; Heinig, U.; Jansen, M.A.K.; Oyama, T.; Pasaribu, B.; Schubert, I.; Sorrels, S.; et al. Return of the Lemnaceae: Duckweed as a model plant system in the genomics and postgenomics era. Plant Cell 2021, 33, 3207–3234.
- Sree, K.S.; Sudakaran, S.; Appenroth, K.-J. How fast can angiosperms grow? Species and clonal diversity of growth rates in the genus Wolffia (Lemnaceae). Acta Physiol. Plant. 2015, 37, 204.
- Ziegler, P.; Adelmann, K.; Zimmer, S.; Schmidt, C.; Appenroth, K.-J. Relative In Vitro growth rates of duckweeds (Lemnaceae)—The most rapidly growing higher plants. Plant Biol. 2015, 17, 33–41.
- Appenroth, K.-J.; Sree, K.S.; Böhm, V.; Hammann, S.; Vetter, W.; Leiterer, M.; Jahreis, G. Nutritional value of duckweeds (Lemnaceae) as human food. Food Chem. 2017, 217, 266–273.
- Skillicorn, P.; Spira, W.; Journey, W. Duckweed Aquaculture: A New Aquatic Farming System for Developing Countries; The World Bank: Washington, DC, USA, 1993.
- Körner, S.; Vermaat, J.E. The relative importance of Lemna gibba L.; bacteria and algae for the nitrogen and phosphorus removal in duckweed-covered domestic wastewater. Water Res. 1998, 32, 3651–3661.
- Xu, J.; Shen, G. Effects of harvest regime and water depth on nutrient recovery from swine wastewater by growing Spirodela oligorrhiza. Water Environ. Res. 2011, 83, 2049–2056.
- Ekperusi, A.O.; Sikoki, F.D.; Nwachukwu, E.O. Application of common duckweed (Lemna minor) in phytoremediation of chemicals in the environment: State and future perspective. Chemosphere 2019, 223, 285–309.
- Roman, B.; Brennan, R.A. Coupling ecological wastewater treatment with the production of livestock feed and irrigation water provides net benefits to human health and the environment: A life cycle assessment. J. Environ. Manag. 2021, 288, 112361.
- Yao, Y.; Zhang, M.; Tian, Y.; Zhao, M.; Zhang, B.; Zhao, M.; Zeng, K.; Yin, B. Duckweed (Spirodela polyrhiza) as green manure for increasing yield and reducing nitrogen loss in rice production. Field Crops Res. 2017, 214, 273–282.
- Yuan, J.; Xu, K. Effects of simulated microgravity on the performance of the duckweeds Lemna aequinoctialis and Wolffia globosa. Aquat. Bot. 2017, 137, 65–71.
- Haynes, R.R.; Les, D.H. Alismatales (Water Plantains). In eLS; Wiley: Hoboken, NJ, USA, 2005.
- Ortiz, O.O.; Ibáñez, A.; Trujillo-Trujillo, E.; Croat, T.B. The emergent macrophyte Montrichardia linifera (Arruda) Schott (Alismatales: Araceae), a rekindled old friend from the Pacific Slope of lower Central America and western Colombia. Nord. J. Bot. 2020, 38, 1–10.
- Bonora, A.; Pancaldi, S.; Gualandri, R.; Fasulo, M.P. Carotenoid and ultrastructure variations is plastids of Arum italicum Miller fruit during maturation and ripening. J. Exp. Bot. 2000, 51, 873–884.
- Demmig-Adams, B.; Stewart, J.J.; Adams, W.W., III. Photoprotection and the trade-off between abiotic and biotic defense. In Non-Photochemical Quenching and Energy Dissipation in Plants, Advances in Photosynthesis and Respiration; Demmig-Adams, B., Garab, G., Adams, W.W., III, Govindjee, U., Eds.; Springer: Dordrecht, The Netherlands, 2014; Volume 40, pp. 631–643.
- Demmig-Adams, B.; Stewart, J.J.; López-Pozo, M.; Polutchko, S.K.; Adams, W.W., III. Zeaxanthin, a molecule for photoprotection in many different environments. Molecules 2020, 25, 5825.
- Demmig-Adams, B.; Ebbert, V.; Mellman, D.L.; Mueh, K.E.; Schaffer, L.; Funk, C.; Zarter, C.R.; Adamska, I.; Jansson, S.; Adams, W.W., III. Modulation of PsbS and flexible vs sustained energy dissipation by light environment in different species. Physiol. Plant. 2006, 127, 670–680.
- Demmig-Adams, B.; Adams, W.W., III; Barker, D.H.; Logan, B.A.; Verhoeven, A.S.; Bowling, D.R. Using chlorophyll fluorescence to assess the allocation of absorbed light to thermal dissipation of excess excitation. Physiol. Plant. 1996, 98, 253–264.
- Demmig-Adams, B.; Adams, W.W., III. Capacity for energy dissipation in the pigment bed in leaves with different xanthophyll cycle pools. Funct. Plant Biol. 1994, 21, 575–588.
- Polutchko, S.K.; Stewart, J.J.; Demmig-Adams, B. Integrative view of the nutrition of the eye. In Nutraceuticals and Functional Foods in Human Health and Disease Prevention; Bagchi, D., Preuss, H.G., Swaroop, A., Eds.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2015; pp. 407–417.
- Demmig-Adams, B.; López-Pozo, M.; Stewart, J.J.; Adams, W.W., III. Zeaxanthin and lutein: Photoprotectors, anti-inflammatories, and brain food. Molecules 2020, 25, 3607.
- Polutchko, S.K.; Glime, G.N.E.; Demmig-Adams, B. Synergistic action of membrane-bound and water-soluble antioxidants in neuroprotection. Molecules 2021, 26, 5385.
- Stewart, J.J.; Adams, W.W., III; Escobar, C.M.; López-Pozo, M.; Demmig-Adams, B. Growth and essential carotenoid micronutrients in Lemna gibba as a function of growth light intensity. Front. Plant Sci. 2020, 11, 480.
- Stewart, J.J.; Adams, W.W., III; López-Pozo, M.; Doherty Garcia, N.; McNamara, M.; Escobar, C.M.; Demmig-Adams, B. Features of the duckweed Lemna that support rapid growth under extremes of light intensity. Cells 2021, 10, 1481.
- Demmig-Adams, B.; Adams, W.W. III. Carotenoid composition in sun and shade leaves of plants with different life forms. Plant Cell Environ. 1992, 15, 411–419.
- Zarter, C.R.; Adams, W.W., III; Ebbert, V.; Adamska, I.; Jansson, S.; Demmig-Adams, B. Winter acclimation of PsbS and related proteins in the evergreen Arctostaphylos uva-ursi as influenced by altitude and light environment. Plant Cell Environ. 2006, 29, 869–878.
- Zarter, C.R.; Adams, W.W., III; Ebbert, V.; Cuthbertson, D.J.; Adamska, I.; Demmig-Adams, B. Winter down-regulation of intrinsic photosynthetic capacity coupled with up-regulation of Elip-like proteins and persistent energy dissipation in a subalpine forest. New Phytol. 2006, 172, 272–282.
- Verhoeven, A.S.; Demmig-Adams, B.; Adams, W.W., III. Enhanced employment of the xanthophyll cycle and thermal energy dissipation in spinach exposed to high light and N stress. Plant Physiol. 1997, 113, 817–824.
- Logan, B.A.; Demmig-Adams, B.; Rosenstiel, T.N.; Adams, W.W., III. Effect of nitrogen limitation on foliar antioxidants in relationship to other metabolic characteristics. Planta 1999, 209, 213–220.
- Demmig-Adams, B.; Stewart, J.J.; Adams, W.W., III. Environmental regulation of intrinsic photosynthetic capacity: An integrated view. Curr. Opin. Plant Biol. 2017, 37, 34–41.
- Terashima, I. Productive structure of a leaf. In Photosynthesis; Briggs, W.R., Ed.; Alan R. Liss: New York, NY, USA, 1989; pp. 207–226.
- Adams, W.W., III; Demmig-Adams, B.; Barker, D.H.; Kiley, S. Carotenoids and photosystem II characteristics of upper and lower halves of leaves acclimated to high light. Aust. J. Plant Physiol. 1996, 23, 669–677.
- Havaux, M.; Niyogi, K.K. The violaxanthin cycle protects plants from photooxidative damage by more than one mechanism. Proc. Natl. Acad. Sci. USA 1999, 96, 8762–8767.
- Havaux, M.; García-Plazaola, J.I. Beyond non-photochemical fluorescence quenching: The overlapping antioxidant functions of zeaxanthin and tocopherols. In Non-Photochemical Quenching and Energy Dissipation in Plants, Advances in Photosynthesis and Respiration; Demmig-Adams, B., Garab, G., Adams, W.W., III, Govindjee, U., Eds.; Springer: Dordrecht, The Netherlands, 2014; Volume 40, pp. 583–603.
- Nilkens, M.; Kress, E.; Lambrev, P.; Miloslavina, Y.; Müller, M.; Holzwarth, A.R.; Jahns, P. Identification of a slowly inducible zeaxanthin-dependent component of non-photochemical quenching of chlorophyll fluorescence generated under steady-state conditions in Arabidopsis. Biochim. Biophys. Acta 2010, 1797, 466–475.
- Muztar, J.A.; Slinger, S.J.; Burton, J.H. Chemical composition of aquatic macrophytes I. Investigation of organic constituents and nutritional potential. Can. J. Plant Sci. 1978, 58, 829–841.
- Rice, S.K. Patterns of allocation and growth in aquatic Sphagnum species. Can. J. Bot. 1995, 73, 349–359.
- Swapna, M.M.; Prakashkumar, R.; Anoop, K.P.; Manju, C.N.; Rajith, N.P. A review on the medicinal and edible aspects of aquatic and wetland plants of India. J. Med. Plant Res. 2011, 5, 7163–7176.
- Huang, M.; Shan, S.; Zhou, X.; Chen, J.; Cao, F.; Jiang, L.; Zou, Y. Leaf photosynthetic performance related to higher radiation use efficiency and grain yield in hybrid rice. Field Crops Res. 2016, 193, 87–93.
- Garbulsky, M.F.; Peñuelas, J.; Gamon, J.; Inoue, Y.; Filella, I. The photochemical reflectance index (PRI) and the remote sensing of leaf, canopy and ecosystem radiation use efficiencies: A review and meta-analysis. Remote Sens. Environ. 2011, 115, 281–297.
- Porcar-Castell, A.; Tyystjärvi, E.; Atherton, J.; van der Tol, C.; Flexas, J.; Pfündel, E.E.; Moreno, J.; Frankenberg, C.; Berry, J.A. Linking chlorophyll a fluorescence to photosynthesis for remote sensing applications: Mechanisms and challenges. J. Exp. Bot. 2014, 65, 4065–4095.
- Thouvenot, L.; Haury, J.; Thiebaut, G. A success story: Water primroses, aquatic plant pests. Aquat. Conserv. Mar. Freshw. Ecosyst. 2013, 23, 790–803.
- Chen, F.; Liu, X.; Yu, C.; Chen, Y.; Tang, H.; Zhang, L. Water lilies as emerging models for Darwin’s abominable mystery. Hortic. Res. 2017, 4, 17051.
- Michael, T.P.; Ernst, E.; Hartwick, N.; Chu, P.; Bryant, D.; Gilbert, S.; Ortleb, S.; Baggs, E.L.; Sree, K.S.; Appenroth, K.J.; et al. Genome and time-of-day transcriptome of Wolffia australiana link morphological minimization with gene loss and less growth control. Genome Res. 2021, 31, 225–238.
- Hsiao, T.C. Plant responses to water stress. Annu. Rev. Plant Physiol. 1973, 24, 519–570.
- Dolferus, R. To grow or not to grow: A stressful decision for plants. Plant Sci. 2014, 229, 247–261.
- Patten, D.T. Productivity and production efficiency of an upper Sonoran Desert ephemeral community. Am. J. Bot. 1978, 65, 891–895.
- Parray, J.A.; Mir, M.Y.; Shameem, N. Stress management: Sustainable approach towards resilient agriculture. In Sustainable Agriculture: Biotechniques in Plant Biology; Parray, J.A., Mir, M.Y., Shameem, N., Eds.; Springer: Singapore, 2019; pp. 231–270.
- Beena, R.; Kirubakaran, S.; Nithya, N.; Manickavelu, A.; Sah, R.P.; Abida, P.S.; Sreekumar, J.; Jaslam, P.M.; Rejeth, R.; Jayalekshmy, V.G.; et al. Association mapping of drought tolerance and agronomic traits in rice (Oryza sativa L.) landraces. BMC Plant Biol. 2021, 21, 484.
- Nilsen, E.T.; Sharifi, M.R.; Rundel, P.W.; Jarrell, W.M.; Virginia, R.A. Diurnal and seasonal water relations of the desert phreatophyte Prosopis glandulosa (honey mesquite) in the Sonoran Desert of California. Ecology 1983, 64, 1381–1393.
- Nilsen, E.T.; Sharifi, M.R.; Rundel, P.W. Comparative water relations of phreatophytes in the Sonoran Desert of California. Ecology 1984, 65, 767–778.
- Sala, A.; Smith, S.D.; Devitt, D.A. Water use by Tamarix ramosissima and associated phreatophytes in a Mojave Desert floodplain. Ecol. Appl. 1996, 6, 888–898.
- Hultine, K.R.; Williams, D.G.; Burgess, S.S.O.; Keefer, T.O. Contrasting patterns of hydraulic redistribution in three desert phreatophytes. Oecologia 2003, 135, 167–175.
- Hultine, K.R.; Koepke, D.F.; Pockman, W.T.; Fravolini, A.; Sperry, J.S.; Williams, D.G. Influence of soil texture on hydraulic properties and water relations of a dominant warm-desert phreatophyte. Tree Physiol. 2006, 26, 313–323.
- Guo, J.S.; Gear, L.; Hultine, K.R.; Koch, G.W.; Ogle, K. Non-structural carbohydrate dynamics associated with antecedent stem water potential and air temperature in a dominant desert shrub. Plant Cell Environ. 2020, 43, 1467–1483.
- Carvajal, D.E.; Loayza, A.P.; Rios, R.S.; Gianoli, E.; Squeo, F.A. Population variation in drought-resistance strategies in a desert shrub along an aridity gradient: Interplay between phenotypic plasticity and ecotypic differentiation. Perspect. Plant Ecol. Evol. Syst. 2017, 29, 12–19.
- Myers, S.S.; Zanobetti, A.; Kloog, I.; Huybers, P.; Leakey, A.D.B.; Bloom, A.J.; Carlisle, E.; Dietterich, L.H.; Fitzgerald, G.; Hasegawa, T.; et al. Increasing CO2 threatens human nutrition. Nature 2014, 510, 139–142.
- Demmig-Adams, B.; Polutchko, S.K.; Zenir, M.C.; Fourounjian, P.; Stewart, J.J.; López-Pozo, M.; Adams, W.W., III. Intersections: Photosynthesis, abiotic stress, and the plant microbiome. Photosynthetica, 2022; in press.
- Körner, C. Paradigm shift in plant growth control. Curr. Opin. Plant Biol. 2015, 25, 107–114.
- Krapp, A.; Stitt, M. An evaluation of direct and indirect mechanisms for the “sink-regulation” of photosynthesis in spinach: Changes in gas exchange, carbohydrates, metabolites, enzyme activities and steady-state transcript levels after cold-girdling source leaves. Planta 1995, 195, 313–323.
- Tausz-Posch, S.; Tausz, M.; Bourgault, M. Elevated effects on crops: Advances in understanding acclimation, nitrogen dynamics and interactions with drought and other organisms. Plant Biol. 2020, 22, 38–51.
- Huang, H.; Ullah, F.; Zhou, D.X.; Yi, M.; Zhao, Y. Mechanisms of ROS regulation of plant development and stress responses. Front. Plant Sci. 2019, 10, 800.
- Foyer, C.H.; Noctor, G. Stress-triggered redox signalling: What’s in pROSpect? Plant Cell Environ. 2016, 39, 951–964.
- Foyer, C.H.; Noctor, G. Redox homeostasis and signaling in a higher-CO2 world. Annu. Rev. Plant Biol. 2020, 71, 157–182.
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Parvin, K.; Bhuiyan, T.F.; Anee, T.I.; Nahar, K.; Hossen, M.S.; Zulfiqar, F.; Alam, M.M.; Fujita, M. Regulation of ros metabolism in plants under environmental stress: A review of recent experimental evidence. Int. J. Mol. Sci. 2020, 21, 8695.
- Padhan, B.K.; Sathee, L.; Meena, H.S.; Adavi, S.B.; Jha, S.K.; Chinnusamy, V. CO2 Elevation accelerates phenology and alters carbon/nitrogen metabolism vis-à-vis ROS abundance in bread wheat. Front. Plant Sci. 2020, 11, 1061.
- Yin, Y.; Yu, C.; Yu, L.; Zhao, J.; Sun, C.; Ma, Y.; Zhou, G. The influence of light intensity and photoperiod on duckweed biomass and starch accumulation for bioethanol production. Bioresour. Technol. 2015, 187, 84–90.
- Paul, M.J.; Foyer, C.H. Sink regulation of photosynthesis. J. Exp. Bot. 2001, 52, 1383–1400.
- Martindale, W.; Bowes, G. The effects of irradiance and CO2 on the activity and activation of ribulose-1,5-bisphosphate carboxylase/oxygenase in the aquatic plant Spirodela polyrhiza. J. Exp. Bot. 1996, 47, 781–784.
- Dall’Osto, L.; Lico, C.; Alric, J.; Giuliano, G.; Havaux, M.; Bassi, R. Lutein is needed for efficient chlorophyll triplet quenching in the major LHCII antenna complex of higher plants and effective photoprotection in vivo under strong light. BMC Plant Biol. 2006, 6, 32.
- Telfer, A. Singlet oxygen production by PSII under light stress: Mechanism, detection and the protective role of β-carotene. Plant Cell Physiol. 2014, 55, 1216–1223.
- Schenk, R.U.; Hildebrandt, A.C. Medium and techniques for induction and growth of monocotyledonous and dicotyledonous plant cell cultures. Can. J. Bot. 1972, 50, 199–204.
- Paul, M.J.; Driscoll, S.P. Sugar repression of photosynthesis: The role of carbohydrates in signalling nitrogen deficiency through source:sink imbalance. Plant Cell Environ. 1997, 20, 110–116.
- Burnett, A.C.; Rogers, A.; Rees, M.; Osborne, C.P. Nutrient sink limitation constrains growth in two barley species with contrasting growth strategies. Plant Direct 2018, 2, e00094.
- Moore, B.D.; Cheng, S.; Sims, D.; Seemann, J.R. The biochemical and molecular basis for photosynthetic acclimation to elevated atmospheric CO2. Plant Cell Environ. 1999, 22, 567–582.
- Ainsworth, E.A.; Long, S.P. 30 years of free-air carbon dioxide enrichment (FACE): What have we learned about future crop productivity and its potential for adaptation? Glob. Chang. Biol. 2005, 27, 27–49.
- Prins, A.; Kunert, K.; Foyer, C.H. Atmospheric CO2 signalling, cellular redox state and plant growth and development. In Redox Metabolism and Longevity Relationships in Animals and Plants; Foyer, C.H., Faragher, R., Thornalley, P., Eds.; Taylor & Francis: London, UK, 2008; Volume 62, pp. 229–252.
- Dai, N.; Schaffer, A.; Petreikov, M.; Shahak, Y.; Giller, Y.; Ratner, K.; Levine, A.; Granot, D. Overexpression of Arabidopsis hexokinase in tomato plants inhibits growth, reduces photosynthesis, and induces rapid senescence. Plant Cell 1999, 11, 1253–1266.
- Diaz, C.; Purdy, S.; Christ, A.; Morot-Gaudry, J.-F.; Wingler, A.; Masclaux-Daubresse, C. Characterization of markers to determine the extent and variability of leaf senescence in Arabidopsis. A metabolic profiling approach. Plant Physiol. 2005, 138, 898–908.
- Agüera, E.; De la Haba, P. Leaf senescence in response to elevated atmospheric CO2 concentration and low nitrogen supply. Biol. Plant. 2018, 62, 401–408.
- Wingler, A.; Purdy, S.; MacLean, J.A.; Pourtau, N. The role of sugars in integrating environmental signals during the regulation of leaf senescence. J. Exp. Bot. 2005, 57, 391–399.
- Wang, S.; Zhang, Y.; Ju, W.; Chen, J.M.; Ciais, P.; Cescatti, A.; Sardans, J.; Janssens, I.A.; Wu, M.; Berry, J.A.; et al. Recent global decline of CO2 fertilization effects on vegetation photosynthesis. Science 2020, 370, 1295–1300.
- Cheng, J.J.; Stomp, A.-M. Growing duckweed to recover nutrients from wastewaters and for production of fuel ethanol and animal feed. Clean Soil Air Water 2009, 37, 17–26.
- Xu, J.; Zhao, H.; Stomp, A.-M.; Cheng, J.J. The production of duckweed as a source of biofuels. Biofuels 2012, 3, 589–601.
- Tao, X.; Fang, Y.; Xiao, Y.; Jin, Y.L.; Ma, X.R.; Zhao, Y.; He, K.Z.; Zhao, H.; Wang, H.Y. Comparative transcriptome analysis to investigate the high starch accumulation of duckweed (Landoltia punctata) under nutrient starvation. Biotechnol. Biofuels 2013, 6, 72.
- Liu, Y.; Wang, X.; Fang, Y.; Huang, M.; Chen, X.; Zhang, Y.; Zhao, H. The effects of photoperiod and nutrition on duckweed (Landoltia punctata) growth and starch accumulation. Ind. Crops Prod. 2018, 115, 243–249.
- Appenroth, K.-J.; Sree, K.S.; Fakhoorian, T.; Lam, E. Resurgence of duckweed research and applications: Report from the 3rd International Duckweed Conference. Plant Mol. Biol. 2015, 89, 647–654.
- Adavi, S.B.; Sathee, L. Elevated CO2 differentially regulates root nitrate transporter kinetics in a genotype and nitrate dose-dependent manner. Plant Sci. 2021, 305, 110807.
- Cedergreen, N.; Madsen, T.V. Nitrogen uptake by the floating macrophyte Lemna minor. New Phytol. 2002, 155, 285–292.
- Cedergreen, N.; Madsen, T.V. Light regulation of root and leaf NO3−uptake and reduction in the floating macrophyte Lemna minor. New Phytol. 2004, 161, 449–457.
- Yu, C.; Zhao, X.; Qi, G.; Bai, Z.; Wang, Y.; Wang, S.; Ma, Y.; Liu, Q.; Hu, R.; Zhou, G. Integrated analysis of transcriptome and metabolites reveals an essential role of metabolic flux in starch accumulation under nitrogen starvation in duckweed. Biotechnol. Biofuels 2017, 10, 167.
- Zhao, Z.; Shi, H.; Liu, Y.; Zhao, H.; Su, H.; Wang, M.; Zhao, Y. The influence of duckweed species diversity on biomass productivity and nutrient removal efficiency in swine wastewater, Bioresour. Technol. 2014, 167, 383–389.
- Mohedano, R.A.; Costa, R.H.R.; Tavares, F.A.; Belli Filho, P. High nutrient removal rate from swine wastes and protein biomass production by full-scale duckweed ponds. Bioresour. Technol. 2012, 112, 98–104.
- Boyd, C.E. Fresh-water plants: A potential source of protein. Econ. Bot. 1968, 22, 359–368.
- Dewanji, A.; Chanda, S.; Si, L.; Barik, S.; Matai, S. Extractability and nutritional value of leaf protein from tropical aquatic plants. Plant Foods Hum. Nutr. 1997, 50, 349–357.
- Rai, U.N.; Sinha, S. Distribution of metals in aquatic edible plants: Trapa natans (Roxb.) Makino and Ipomoea aquatica Forsk. Environ. Monit. Assess. 2001, 70, 241–252.
- Zeng, J.; Sheng, H.; Liu, Y.; Wang, Y.; Wang, Y.; Kang, H.; Fan, X.; Sha, L.; Yuan, S.; Zhou, Y. High nitrogen supply induces physiological responsiveness to long photoperiod in barley. Front. Plant Sci. 2017, 8, 569.
- Du, S.; Zhang, Y.; Lin, X.; Wang, Y.; Tang, C. Regulation of nitrate reductase by nitric oxide in Chinese cabbage pakchoi (Brassica chinensis L.). Plant Cell Environ. 2008, 31, 195–204.
- Bian, Z.; Wang, Y.; Zhang, X.; Li, T.; Grundy, S.; Yang, Q.; Cheng, R. A review of environment effects on nitrate accumulation in leafy vegetables grown in controlled environments. Foods 2020, 9, 732.
- Shin, R.; Schachtman, D.P. Hydrogen peroxide mediates plant root cell response to nutrient deprivation. Proc. Natl. Acad. Sci. USA 2004, 101, 8827–8832.
- Kim, M.J.; Ciani, S.; Schachtman, D.P. A peroxidase contributes to ros production during Arabidopsis root response to potassium deficiency. Mol. Plant 2010, 3, 420–427.
- Wu, F.; Sun, X.; Hu, X.; Zou, B.; Lin, N.; Lin, J.; Ji, K. Response of nitrogen metabolism in masson pine needles to elevated CO2. Forests 2020, 11, 390.
- Adavi, S.B.; Sathee, L. Elevated CO2 alters tissue balance of nitrogen metabolism and downregulates nitrogen assimilation and signalling gene expression in wheat seedlings receiving high nitrate supply. Protoplasma 2020, 258, 219–233.
- Queval, G.; Issakidis-Bourguet, E.; Hoeberichts, F.A.; Vandorpe, M.; Gakière, B.; Vanacker, H.; Miginiac-Maslow, M.; Van Breusegem, F.; Noctor, G. Conditional oxidative stress responses in the Arabidopsis photorespiratory mutant cat2 demonstrate that redox state is a key modulator of daylength-dependent gene expression, and define photoperiod as a crucial factor in the regulation of H2O2-induced cell death. Plant J. 2007, 52, 640–657.
- Krasensky-Wrzaczek, J.; Kangasjärvi, J. The role of reactive oxygen species in the integration of temperature and light signals. J. Exp. Bot. 2018, 69, 3347–3358.
- Begara-Morales, J.C. Nitric oxide signalling in a CO2-enriched environment. J. Exp. Bot. 2016, 67, 560–561.
- Bloom, A.J. As carbon dioxide rises, food quality will decline without careful nitrogen management. Calif. Agric. 2009, 63, 67–72.
- de Sousa Leite, T.; Monteiro, F.A. Partial replacement of nitrate by ammonium increases photosynthesis and reduces oxidative stress in tanzania guinea grass exposed to cadmium. Ecotoxicol. Environ. Saf. 2019, 174, 592–600.
- Britto, D.T.; Kronzucker, H.J. NH4+ toxicity in higher plants: A critical review. J. Plant Physiol. 2002, 159, 567–584.