The Colorado potato beetle (CPB) is one of the most important potato pest worldwide. It is native to U.S. but during the 20th century it has dispersed through Europe, Asia and western China. It continues to expand in an east and southeast direction. Damages are caused by larvae and adults. Their feeding on potato plant leaves can cause complete defoliation and lead to a large yield loss. After the long period of using only chemical control measures, the emergence of resistance increased and some new and different methods come to the fore. The main focus of this review is on new approaches to the old CPB control problem. We describe the use of Bacillus thuringiensis and RNA interference (RNAi) as possible solutions for the future in CPB management. RNAi has proven successful in controlling many pests and shows great potential for CPB control. Better understanding of the mechanisms that affect efficiency will enable the development of this technology and boost potential of RNAi to become part of integrated plant protection in the future. We described also the possibility of using single nucleotide polymorphisms (SNPs) as a way to go deeper into our understanding of resistance and how it influences genotypes.
- Colorado potato beetle
- resistance problem
- control strategies
- GM potato
- RNAi
- SNPs
1. Introduction
Colorado Potato Beetle—A Global Pest of Potato Production
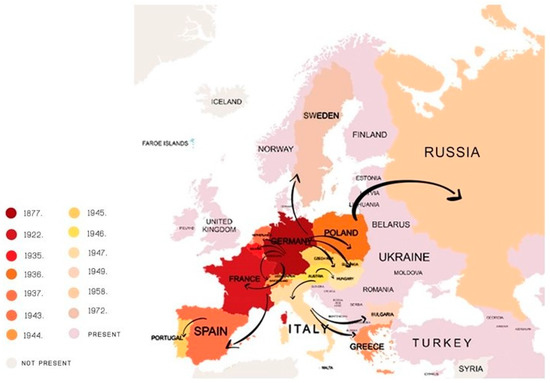

2. Bacillus thuringiensis (Bt) in the Fight to Control Colorado Potato Beetle
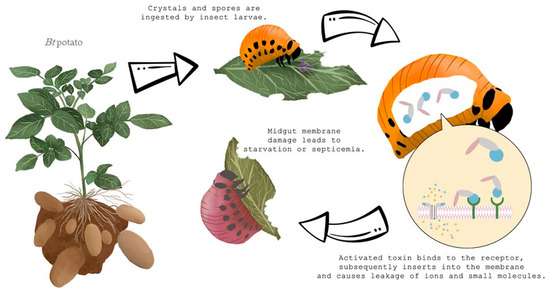
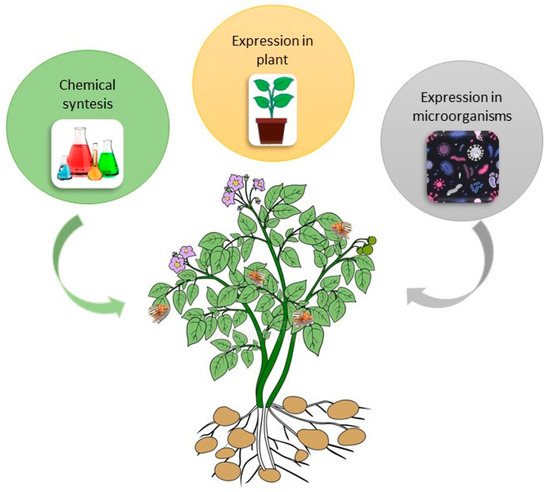
2.1. Bt Potato Development
2.2. Why Bt Potato Did Not Sustain on Market
3. Sources of Host-Plant Resistance
4. New Approaches to Colorado Potato Beetle Management
4.1. RNA Interference (RNAi)
4.2. RNAi in Colorado Potato Beetle Control Management
| Target Gene | Annotation | Reference |
|---|---|---|
| VATPase, A, B, E | Vacuolar ATP synthase subunit | [61,65] |
| Sec23 | Protein transport protein sec23 | [65] |
| COPβ | Coatomer β-subunit | [65] |
| Actin | β-Actin | [65] |
| Prohibitin | Prohibitin protein | [70] |
| SAHase | S-adenosyl-L-homocysteine hydrolase | [66] |
| FTZ-F1 | Nuclear receptor type transcription factor that responses to 20-hydroxyecdysone |
[74] |
| shd | Ecdysone 20-monooxygenase | [69] |
| NAT1 | Nutrient amino acid transporter | [75] |
| Actin | β-Actin | [76] |
| JHEH | Juvenile hormone epoxide hydrolase | [77] |
| alt | Alanine aminotransferase | [71] |
| p5cdh | Δ1-pyrroline-5-carboxylate dehydrogenase | [72] |
| HR3 | Nuclear receptor that early-late responses to 20-Hydroxyecdysone |
[78] |
| UAP | Uridine diphosphate N-acetylglucosamine pyrophosphorylase |
[79] |
| ChS | Chitin synthase | [80] |
| TPS and TREs | Trehalose biosynthesis and degradation | [81] |
| E75 | Ecdysone-induced protein 75 | [82] |
| JHAMT | Juvenile hormone acid methyltransferase | [83] |
| ILP2 | Putative insulin-like peptide | [84] |
| HR4 | ecdysteroidogenesis and mediates 20-hydroxyecdysone signaling during larval-pupal metamorphosis | [85] |
| CncC | Xenobiotic transcription factor | [69] |
| EcR | Ecdysone receptor | [73] |
| Mesh | gut-membrane-associated protein | [86] |
4.3. RNAi Based Products in Wide Use
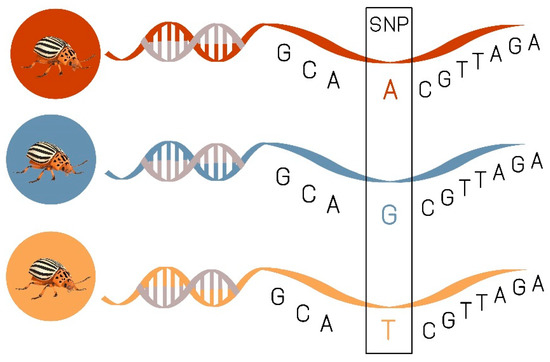
5. Genetic Tools in Colorado Potato Beetle Management
Single Nucleotide Polymorphism (SNPs) as Prospective Tool in CPB Resistance Management
6. Conclusions
This entry is adapted from the peer-reviewed paper 10.3390/insects11090581