Liver fibrosis is a wound healing response that degenerates, and is characterized by excessive accumulation of extracellular matrix (ECM) components that form scar tissue. Liver transplantation is often needed as a course of treatment for patients in critical conditions, but limitations associated with transplantation prompted the continuous search for alternative therapeutic strategies. Cell therapy with stem cells has emerged as an attractive option in order to stimulate tissue regeneration and liver repair. Transplanted mesenchymal stem cells (MSCs) could trans-differentiate into hepatocyte-like cells and, moreover, show anti-fibrotic and immunomodulatory effects.
Introduction
Liver fibrosis is a wound healing response that degenerates, and is characterized by excessive accumulation of extracellular matrix (ECM) components that form scar tissue [
1]. There are many causes for liver fibrosis, such as alcohol abuse, non-alcoholic steatohepatitis (NASH), viral or autoimmune hepatitis, non-alcoholic fatty liver disease (NAFLD), ischemic injury, and congenital syndromes [
2,
3]. Liver fibrosis, left untreated, can progress towards irreversible end-stages of liver failure, liver cirrhosis and hepatocellular carcinoma [
4]. Most frequently, either removing the cause or the use of antifibrotic drugs would not be completely effective in treating the disease, and liver transplantation is often recommended [
5].
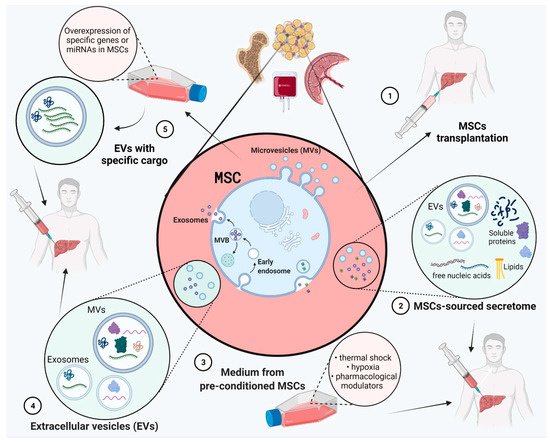
Cell therapies have emerged as an alternative to liver transplantation in order to induce tissue regeneration (
Figure 1). Numerous cell sources have been investigated for their regenerative potential, either hepatocytes [
6], or different types of stem cells [
7,
8]. There are two major categories of stem cells, embryonic stem cells (ESCs) and adult stem cells (ASCs). Two sources of pluripotent stem cells that can differentiate into hepatocyte-like cells are ESCs, derived from embryos [
9,
10,
11], and induced pluripotent stem cells (iPSCs) [
12]. ASCs are multipotent stem cells and have a more limited cell differentiation potential than ESCs. ASCs considered for liver regeneration are liver stem cells (LSCs) [
13,
14] and mesenchymal stem cells (MSCs) [
15,
16] from different tissue sources, such as bone marrow (BM-MSCs) [
17], adipose tissue (ADSCs) [
18,
19,
20], umbilical cords (UC-SCs), peripheral blood, cartilage, etc. MSCs show low immunogenicity, have a self-renewal ability, are easily obtainable, and can be used without ethical issues, which make them the perfect candidate for liver regeneration [
21]. They can migrate to injury sites as a response to cellular damage signals, and they have been shown to promote other cells’ migration to liver sites, differentiate in hepatocyte-like cells and participate in liver regeneration via paracrine mechanisms [
22,
23,
24].
Figure 1. Liver fibrosis therapies based on mesenchymal stem cells (MSCs). MSCs can be isolated from many organs and tissues such as bone marrow, adipose tissue, peripheral blood, placenta, umbilical cord etc. MSC-based therapy strategies: (1) MSCs transplantation directly to the patient; (2) Use of MSC-sourced secretome (soluble molecules and extracellular vesicles (EVs)); (3) Use of medium from pre-conditioned MSCs (physical or environmental shock and pharmacological modulators); (4) Use of MSC-sourced EVs; (5) Use of MSC-sourced EVs with upregulated expression of genes or miRNAs. Figure created with BioRender.com on 6 December 2021.
Cell therapies based on MSCs could help liver regeneration directly, by hepatogenic differentiation or cell migration to liver sites, or indirectly, in a paracrine manner via its secretome [
23]. MSC-sourced secretome contains many soluble molecules and extracellular vesicles (EVs) that direct tissue repair and regeneration [
9,
25]. Cell based therapies come with a number of limitations and risks, and cell-free strategies, such as MSC-sourced secretome, could represent an improved alternative. The use of secretome rather than cells avoids the risks associated with tumorigenesis or potential differentiation into pro-fibrotic cells [
26,
27]. MSC-sourced secretome can be stored over long periods of time without toxic cryopreservative agents and also, it could be produced in large quantities over a short period of time with low costs. Moreover, the content of MSC-sourced secretome could be modified for specific desired therapeutic effects [
27]. Cell-based therapies face an additional challenge in regard to its method of transplantation to patients, as such, an effective method has not yet been identified, and cells may end up in other organs [
7].
Composition of MSCs’ Secretome (Derived Soluble Factors CM/EV)
MSCs secrete a number of molecules, such as soluble proteins, free nucleic acids, lipids and EVs, all accounting for MSCs’ secretome [
9,
27,
48]. MSCs can be isolated from many types of tissues and organs, and even though they share many characteristics and show regenerative potential, their secretome content may vary based on their origin. Differences in MSC-sourced secretome have been pointed out between ADSCs, BM-MSCs and UC-SCs [
10,
12,
49]. All types of MSCs secrete factors such as cytokines, chemokines, growth factors, and anti-inflammatory factors. Overall, it was proved that MSC-sourced secretome shows immunomodulatory, anti-inflammatory and anti-apoptotic activity, regulates angiogenesis, stimulates wound healing and tissue repair, and has antitumor and anti-microbial effects [
27]. However, the exact mechanisms through which MSC-sourced secretome exerts its role are not fully understood, and the exact molecules responsible for each effect are not specifically correlated.
MSCs secrete not only a number of anti-inflammatory, but also pro-inflammatory cytokines which mediate its immunomodulatory effect. The secretome contains anti-inflammatory cytokines, such as tumor necrosis factor β1 (TNFβ1), interleukin (IL) 13, IL18 binding protein (IL18BP), ciliary neutrophic factor (CNTF), neurotrophin 3 (NT-3) factor, IL10, IL12p70, IL17E, IL27, IL1. MSCs also secrete pro-inflammatory cytokines, such as IL1b, IL6, IL8 and IL9 [
27] and the pleiotropic cytokine transforming growth factor β (TGF-β) [
49]. Moreover, MSCs secrete growth factors with regenerative potential, such as vascular endothelial growth factor (VEGF), hepatocyte growth factor-1 (HGF-1), leukemia inhibitory factor (LIF) [
49], and keratinocyte growth factor (KGF) [
50]. HGF shows anti-fibrotic properties, and can induce apoptosis of activated HSCs and promote hepatocyte proliferation [
51,
52].
MSC-sourced secretome contains different types of EVs: exosomes (exo), microvesicles (MVs) and apoptotic bodies, classified based on their size and biogenesis, but not on other specific markers [
26,
53]. Recent studies suggest that MSC-derived EVs, and especially exosomes, play an important role in the therapeutic effect of MSCs through paracrine mechanism. They are easily obtainable through filtering and ultracentrifugation protocols [
27,
54]. EVs are small particles originated from the plasma membrane, which are secreted from cells, either as part of a disposal mechanisms or as communication mediators between cells [
55]. Exosomes are small spherical bilipid membrane vesicles (40–150 nm) which emerge from fusion with early endosomes and form multivesicular endosomes (MVEs). These will fuse with the cell membrane and exosomes will be expelled from the cell [
56]. MVs are bigger vesicles (100–1000 nm) formed from direct outward budding of the plasma membrane [
26].
Apoptotic bodies (1–5 µm) are formed during the self-destructive actions of a cell during apoptosis, by the outward budding of the plasma membrane. Opposed to exosomes and MVs, apoptotic bodies contain components of cell degradation from dying cells and undergo phagocytosis by macrophages [
57].
EVs hold an important role in intercellular communication as their cargo includes proteins, lipids, mRNA, microRNA and they can be safely transferred between cells. Their cargo is determined by their tissue source and influences their effects. Moreover, MSCs can be modified in order to manipulate the EVs cargo to contain specific molecules [
26,
58,
59].
Role of MSCs-Sourced Secretome in Liver Regeneration
MSCs in Liver Disease Treatment
The practice of MSC transplantation emerged as a way to improve liver function, in terms of bringing an extra source of cells with differentiation potential that would help speed up the regeneration process. In addition, it was known that stem cells-sourced secretome is rich in molecules also involved in regenerative mechanisms.
MSCs could be induced to differentiate into hepatocyte-like cells that can contribute to liver regeneration [
21,
60]. The source of MSCs greatly influences their differentiation potential and the type of cytokines and growth factors needed to induce it [
61]. There is no standard for the growth factor cocktail, which is different based on the MSCs’ source and between different studies. Most of the studies use fibroblast growth factor (FGF) and epidermal growth factor (EGF) in a primary induction step that stimulates the proliferation of MSCs [
62]. HGF is a pleiotropic cytokine which regulates proliferation, differentiation and migration of MSCs, and it is frequently used in differentiation studies [
63]. Other factors used for differentiation are nicotinamide (NTA) and insulin-transferrin selenium (ITS) which promote the proliferation and survival of primary hepatocytes [
64]. Oncostatin M (OSM) and dexamethasone (Dex) also help with the maturation process of developing hepatocytes [
65].
Effect of MSC-Derived Conditioned Medium (MSC-CM)
The conditioned medium (CM) from MSCs contains all the components of the secretome, and is easily obtainable from cultured MSCs by centrifugation and filtration [
27]. CM was frequently used in studies to evaluate the effects of MSC-sourced secretome in liver regeneration, without the risks associated with cell transplantation.
UC-MSCs were isolated and differentiated in vitro into hepatocyte-like cells, and their secretome was obtained in a study by An et al. [
80]. CM-MSCs, from differentiated and undifferentiated UC-MSCs, were distributed to TGF-β1-activated HSCs and mice with thioacetamide (TAA) and CCl
4-induced liver fibrosis. CM-MSCs inhibited HSCs activation and reduced the expression of fibrotic factors such as α-SMA, collagens, metalloproteinases, TGFβ, and Smad proteins in the TGFβ signaling pathway. One highly expressed protein was identified in the UC-MSCs secretome, milk fat globule-EGF factor 8 (MFGE8), and it was proven, in the study, to show anti-fibrotic action similar to the effect of CM-MSC.
Pre-Treatment of MSCs for Improved Secretome Content
Therapeutic effects of MSCs can be improved by changing the cell culture conditions of MSCs that will further modulate the content of MSCs secretome. Preconditioning can prepare the cells for transplantation in vivo and improve their survival rate and its paracrine effects [
85]. Some pro-survival strategies include methods in which cells are exposed to physical or environmental shocks and pharmacological modulators of targeted molecules. One strategy is a thermal preconditioning method at 42 °C for 1–2 h before transplantation, proven to promote cell survival in vivo [
86]. Next, hypoxic preconditioning mimics the natural in vivo conditions (1–12% in vivo compared to 21% in vitro) and improves MSCs multipotency and self-renewal abilities [
87,
88].
Other cell culture conditions can modulate the immunomodulatory properties of MSCs. LPS is a representative Toll-like receptor 4 (TLR4) agonist and can induce MSCs to exert an immunosuppressive phenotype, necessary in order to reduce liver fibrosis-related inflammation [
89,
90]. In a study by Lee et al. [
91], AD-MSCs were pre-cultured for 24 h with a low dose of lipopolysaccharide (LPS). LPS preconditioning of AD-MSCs upregulated the expression of inflammation-related mediators such as IL-6, TNF-α, HGF, and VEGF. CM from LPS-preconditioned AD-MSCs was injected to partially hepatectomized mice, and liver regeneration was found to be improved compared to the control. It was proposed that pro-regenerative effects of the LPS-preconditioned CM were mediated through IL-6/STAT3 activation.
Effect of EVs
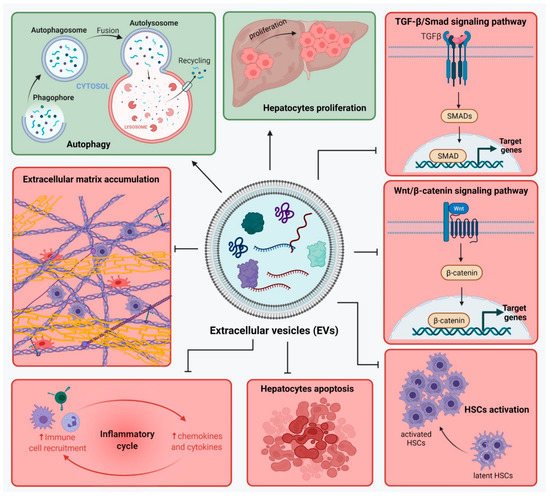
A lot of the recent interest has been focused on the EVs in the MSCs-sourced secretome and their specific role in modulating liver repair. It was shown that MSC-EVs modulate liver regeneration by regulating many pathways involved in liver fibrosis development. These effects were confirmed on EVs from different types of MSCs derived from adipose tissue, bone marrow, umbilical cord blood, placenta or amnion.
MSC-EVs act upon the TGF-β1/Smad signaling pathway [
92,
93] and Wnt/β-catenin pathway [
94,
95], stimulate autophagy, inhibit hepatocyte apoptosis [
96], reduce collagen deposition [
92,
94,
97], inhibit activation of HSCs [
98], modulate the inflammation by reducing levels of inflammatory factors [
99,
100,
101], and inhibit activation of NLRP3 inflammasome [
102] (
Figure 2).
Figure 2. Extracellular vesicles from mesenchymal stem cells (MSC-EVs) modulate liver regeneration by regulating pathways involved in liver fibrosis development. EVs’ content is rich in proteins, lipids, mRNA and microRNA. MSC-EVs may act in two distinct ways: Green—Inhibition of: TGF-β1/Smad signaling pathway, Wnt/β-catenin pathway, hepatic stellate cells (HSCs) activation, hepatocyte apoptosis, inflammatory factors secretion, collagen deposition; Red—Activation of: autophagy and hepatocyte proliferation. Figure created with BioRender.com on 17 November 2021.
Exosomes from MSCs with Specific Overexpressed Cargo Such as miRNAs
An increasing number of studies report enhancing the expression of certain therapeutic genes or miRNAs in MSCs, in order to modulate the cargo of EVs. In addition, the overexpression of specific proteins or miRNAs offers the basis to study their way of action in liver regeneration and how they interact with molecules in signaling pathways.
The expression of miR-122 is reduced in advanced liver diseases, such as cirrhosis, and it is a molecule associated with anti-fibrotic potential [
106,
107]. MiR-122 was overexpressed in AD-MSCs by using a lentivirus (LV)-mediated transfer of pre-miR-122 precursor molecules (LV-miR-122), and exosomes were isolated from the supernatant of the cells using ExoQuick-TC Kit [
104]. Exosomes from miR-122-modified AD-MSCs specifically downregulated, in the LX2 HSC cell line, the expression of miR-122 target genes
P4HA1,
IGF1R and CCNG1, which are involved in HSCs activation and proliferation. Moreover, these exosomes with miR-122 acted in vivo in mice with CCl
4-induced liver fibrosis, and reduced the expression of TGF-β1 and α-SMA, and suppressed the serum levels of HA, P-III-P, ALT, AST and liver hydroxyproline content.
Conclusions
Liver fibrosis is a result of liver injury and an altered wound healing response, which leads to ECM proteins’ accumulation. There are complex interactions between liver cells in liver fibrosis development and many signaling pathways are involved in maintaining the inflammatory state. Therapies for liver fibrosis are constantly under investigation, and the use of strategies based on MSCs show great promise. MSC transplantation showed to improve liver function as it brings a source of cells with differentiation potential towards hepatocyte-like cells. In addition to contributing in restoring the number of hepatocytes in the injured liver, MSC-sourced secretome is rich in molecules involved in regenerative mechanisms by action on liver cells such as hepatocytes, HSCs and macrophages. MSC-secretome acts upon hepatocytes and promotes their survival and proliferation. Especially when MSC-EVs were used, the expression of apoptotic markers was inhibited. In addition, MSCs show immunomodulatory and anti-inflammatory activity though the action of their secretome. They stimulate the production of M2-type macrophages, which will secrete anti-inflammatory molecules. Moreover, MSC-secretome inhibits the infiltration of inflammatory cells and suppresses the expression of inflammatory cytokines in KCs. Activated HSCs represent another important cellular target of the MSC-sourced secretome. Not only does it help reduce their number in the fibrotic liver by stimulating their apoptosis, but it also further inhibits their activation and proliferation, thus helping control the overall pool of HSCs. HSCs are also responsible for the ECM production in liver fibrosis, and MSC-secretome inhibits the production of most ECM proteins and fibrotic markers. In such a manner, MSCs contribute in different ways (direct MSC transplantation, MSC-sourced secretome, CM from pre-treated MSCs, MSC-EVs) to alleviate liver damage and promote tissue regeneration. Cell-free therapies based on MSC-sourced secretome help overcome a number of limitations and risks associated with MSCs transplantation. The use of secretome could mimic the anti-fibrotic effects observed after MSCs transplantation. MSCs secretome is rich not only in cytokines, chemokines, growth factors, and anti-inflammatory factors, but in EVs as well. The use of MSCs-EVs as a treatment for liver fibrosis may be more effective than MSCs therapy, as they can pass through biological barriers and deliver their anti-fibrotic cargo to specific target cells. MSC-EVs were shown to regulate signaling pathways such as TGF-β1/Smad and Wnt/β-catenin, stimulate autophagy and hepatocyte apoptosis, reduce collagen deposition, inhibit activation of HSCs, and modulate the inflammation. Moreover, MSC-EVs cargo can be modified in order to deliver specific proteins of miRNAs with anti-fibrotic properties. So far, these strategies show great promise, but more research is needed to confirm their efficiency and safety, especially for the use of EVs with modified cargo.
This entry is adapted from the peer-reviewed paper 10.3390/ijms222413292