Stink bugs use semiochemicals to communicate over long distances and exchange vibratory signals that are transmitted on plants over shorter distances. These signals are produced by various mechanisms, such as body vibration (tremulation) or drumming on the substrate, and are accompanied by visual, chemical, and mechanical signals and cues when they encounter a mate.
- plant-dwelling insects
- biotremology
- host plants
- sexual communication
- chemical signals
- vibratory signals
1. Communication of Stink Bugs during Calling, Courtship and Rivalry
In many stink bugs, communication begins with the emission of male species-specific pheromones, which attract females from longer distances [66] (Figure 1). Once on the same plant, information exchange between stink bugs takes place via substrate-borne signals [65,66,67]. Insects most frequently use vibrations as the means of communication on plants or other substrates [87]. Studies on stink bugs, and many other insects [13,14,15,89,90] have described various behavioral and neural mechanisms involved in substrate-borne vibratory communication.
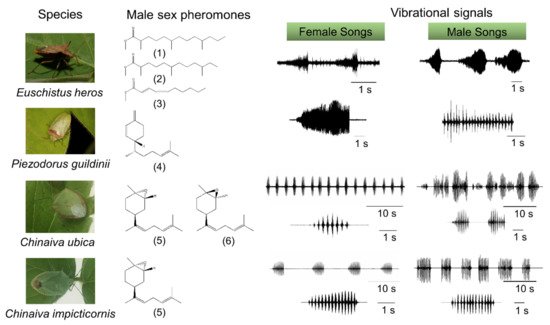
1.1 Vibrational communication
In the southern green stink bug, Nezara viridula (Pentatomidae), a male searches for a mate and emits calling and courtship substrate-borne signals in response to vibratory calls from the stationary female [13,14,15]. In both sexes, these signals are produced by abdominal vibration (AV signals). Many stink bugs form a stereotyped female-male duet that is crucial for mate finding and recognition [64,68,69,70,71,72]. Upon encountering a mate, the vibratory calling phase transitions into a multimodal courtship phase in which AV signals are accompanied by visual and tactile signals and cues, the latter acting through both chemical and mechanical channels [13,14,15]. The exchange of calling and courtship signals between mates is inhibited by male or female rivalry when multiple individuals compete for copulation with the same mate [13,73,74].
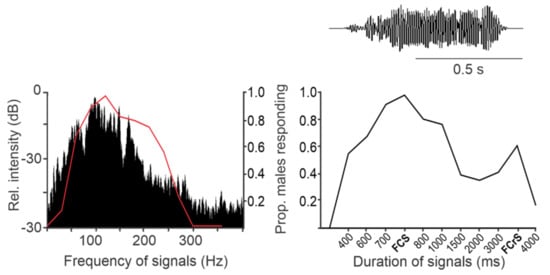
The AV signals have been described in 36 stink bug species as components of calling, courtship, rivalry, and copulation songs [13]. They share a common fundamental (basic) frequency that typically ranges between 90 and 120 Hz (Figure 2), with the extreme lowest and highest values measured to date in N. antennata (50 Hz, female calling song) and Euschistus heros (175 Hz, male courtship song) [75,76]. AV signal amplitudes (expressed in velocity units) range from 0.1 to 1 mm/s when measured on the body of emitter bugs standing on a plant [77].Among the different types of vibratory signals, the AV signals show the greatest diversity of temporal characteristics that determine their species and sex specificity. The temporal pattern of elements of the same song type can differ considerably among populations of the same species, as shown, for example, in N. viridula from Brazil, Florida, Italy, and Slovenia [78]. Accordingly, genetic differences have been described between 11 geographically separated populations of N. viridula from Europe (Slovenia, France, Greece, Italy and Madeira), Japan, Guadalupe, Galapagos, California, Brazil and Botswana [79]. Several playback experiments have confirmed that the duration of song components and the intervals between them mediate crucial information for recognition and directionality [64,71,72,80; Figure 2].
The temporal pattern of calling, courtship, and rivalry songs determines their function in communication [13]. The female calling song is characterized by longer periods of signals with a simple repetitive pattern, enabling directional movement of the male towards the calling mate. The courtship song has a more complex structure, and is supported by signals from other modalities that enhance recognition and motivate the mates to copulate. Male rivalry song of most species studied to date is characterized by a short-term production of rapidly repeated shorter and often frequency-modulated (FM) pulses. Typically, following signal alternation, one of the rivals stops signaling [13,14,15]. Female rivalry is more complex and involves prolonged alternation with the rival signals and modified calling signals [73,74]. In addition, vibrations of the raised wings (buzzing signals) or whole body tremulation have been described as constituents of communication in E. heros, C. impicticornis and C. ubica. These signals are of high amplitude, broad-band nature, and are species- but not sex-specific [59,81]. Buzzing signals usually precede AV signals in the very early phase of communication [81], while whole body tremulation has been observed in the context of aggression (rivalry) and shortly preceding copulation [59]. Moreover, stink bugs produce low-amplitude percussion signals by tapping the substrate with their forelegs. These signals are species- and sex-specific and occur in a variety of contexts, for example, immediately after copulation [13], but the function of these signals has largely not been elucidated.
2. Transmission of Vibratory Signals through Plants
3. The Effect of Vibratory Noise on Communication
Since stink bug vibratory signals are narrow-banded, the sensory system can selectively filter out noise of lower and higher frequencies without affecting signal information. At the same time, species-specificity of signals is not encoded in their frequency domain, consequently implying larger heterospecific overlap [e.g., 98, 104].
Wind and rain represent the main source of noise in the field, characterized by most energy content at very low frequencies (below 30 Hz) [87,101,107,108,109,110,111,112]. These frequencies lie below that characteristic of stink bug AV signals and the peak sensitivity of specialized vibratory receptor organs [117]. Nevertheless, in laboratory experiments, significantly lower copulatory success was observed in E. heros stink bugs exposed to airflow or rain-generated vibratory noise [74]. Additional experiments are needed to test the influence of low-velocity wind on vibrational communication of stink bugs in the field.
Vibratory noise has a number of effects on stink bug communication [74, 113,114, 115]. In N. viridula, for example, exposure of male-female pairs to pure-tone background vibrations with frequency and amplitude characteristics similar to those of conspecific signals had no effect on the expression of male searching behavior. Females, on the other hand, discontinued their calls, reduced the repetition rate of their emissions, or replaced their calls with rival songs. Furthermore, females changed the fundamental frequency of the calling song so to increase its differentiation from the background vibration [113]. Similarly, males and females of E. heros adjusted the repetition rate of their calls and modified their fundamental frequency in response to vibratory noise [98]. These modifications can all be interpreted as the mechanisms for reducing signal masking interference. Rival songs of C. impicticornis, C. ubica, and N. viridula act in a similar way, as they inhibit simultaneous singing by multiple mates [13,14,15]. In contrast, in N. viridula, low-intensity white Gaussian noise presented on a plant along with subthreshold female calling song positively affects male searching behaviour. The intensity-response characteristics of this influence suggest that it is due to the stochastic threshold resonance [115].
This entry is adapted from the peer-reviewed paper 10.3390/insects12121058