C-X-C chemokine ligand 13 (CXCL13) and its receptor, CXCR5, make crucial contributions to this process by triggering intracellular signaling cascades in malignant cells and modulating the sophisticated TME in an autocrine or paracrine fashion. The CXCL13/CXCR5 axis has a dominant role in B cell recruitment and tertiary lymphoid structure formation, which activate immune responses against some tumors. In most cancer types, the CXCL13/CXCR5 axis mediates pro-neoplastic immune reactions by recruiting suppressive immune cells into tumor tissues. Tobacco smoke and haze (smohaze) and the carcinogen benzo(a)pyrene induce the secretion of CXCL13 by lung epithelial cells, which contributes to environmental lung carcinogenesis.
1. Introduction
Chemokines are a family of chemotactic cytokines with small molecular weights (8–14 kDa) [
1]. Chemokines are classified into four groups according to the position of the first two cysteines closest to the amino terminus: C, CC, CXC, and CX3C [
2]. Chemokines exert their functions by binding to their receptors, which are seven-transmembrane guanine-protein-coupled receptors (GPCRs) [
3]. Chemokines have important roles in regulating lymphoid tissue development, immune homeostasis, and inflammatory responses by directing the migration of leukocytes into the injured or infected tissues [
2]. A complex chemokine-chemokine receptor signaling network is critical to the tumor microenvironment (TME), which makes pivotal contributions to tumor cell proliferation, migration, invasion, angiogenesis, and evasion of anti-tumor immunity, facilitating tumor initiation, progression, and metastasis [
4,
5,
6,
7,
8].
2. CXCL13/CXCR5 and Immune Homeostasis
2.1. CXCL13/CXCR5: Genes and Proteins
C-X-C chemokine ligand 13 (CXCL13), also known as B-cell attracting chemokine 1 (BCA-1) or B-lymphocyte chemoattractant (BLC), was originally identified as a homeostatic chemokine to attract B cells, a minority of T cells, and macrophages [
13]. The human
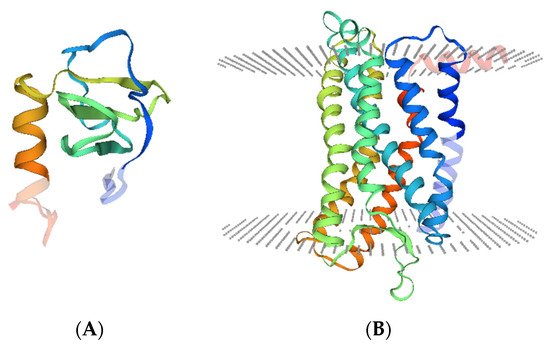
CXCL13 gene localizes on chromosome 4q21 and encodes CXCL13 protein, which has 109 amino acids, a molecular mass of 12,664 Da, and a crystal structure as below (
Figure 1A). The receptor of CXCL13 is the C-X-C chemokine receptor type 5 (CXCR5), which is also named Burkitt’s lymphoma receptor 1 (BLR1) and is defined as a member of the superfamily of seven-transmembrane GPCRs (
Figure 1B). CXCR5 has two transcripts, both localized on the cell membrane [
14], and is expressed by follicular helper T cells (Tfh) [
15], circulating CD4
+ T cells [
16], B cells [
17], CD68
+ macrophages [
18], and tumor cells. Moreover, FANCA-mediated CXCR5 neddylation is involved in targeting the receptor to the cell membrane, and CXCR5 neddylation stimulates cell migration and motility [
19].
Figure 1. Crystal structure of CXCL13 and CXCR5. (
A). Illustration of the CXCL13 monomer (UniProKB-O43927) showing domain hits with deep coloration. (
B). Illustration of the CXCR5 monomer (UniProKB-P32302) showing seven transmembrane helixes and domain hits (deeply colored). Structural models were obtained from SWISS-MODEL (
http://swissmodel.expasy.org/repository/. accessed on 15 April 2021).
2.2. CXCL13/CXCR5 Axis
The precise mechanism of how the CXCR5 receptor responds to CXCL13 and mediates signaling activation has not been fully elucidated. Evidence has demonstrated that CXCR5 interacts with cytosolic and membrane proteins to form heterodimers and heterotrimers, respectively [
20,
21,
22]. CXCR5 couples to cytosolic α, β, and γ subunits of G proteins to form heterotrimeric guanine nucleotide-binding proteins [
20]. After CXCL13 binds to CXCR5, G proteins dissociate from CXCR5, dividing into G
α and G
βγ, which stimulate different downstream molecules and subsequently trigger specific intracellular signal transduction pathways [
20,
23]. The intracellular domains, and probably the transmembrane-spanning domains of CXCR5, are required to activate G proteins [
24,
25]. CXCR5 can also form heterodimers with membrane proteins, such as CXCR4 and Epstein–Barr virus-induced receptor 2 (EBI2) [
20,
22]. The EBI2/CXCR5 heterodimer lowers the affinity of CXCL13 for CXCR5 and reduces the activation of G proteins, potentially contributing to the alteration of the CXCR5 binding pocket by heterodimer formation [
22].
2.3. Physiological Functions of CXCL13/CXCR5
CXCL13 is abundantly expressed on follicular helper T cells (Tfh), follicular dendritic cells (FDCs), and stromal cells in the follicles of secondary lymphoid organs (SLOs) and is essential for the development of the B cell zones of SLOs [
13,
26,
27,
28]. SLOs, which include the spleen, lymph nodes, and Peyer’s patches, coordinate antigen-specific primary immune responses via promoting the interactions between antigen-presenting cells and lymphocytes. CXCR5 is expressed by mature B lymphocytes [
29], a subpopulation of follicular B helper T cells [
30,
31], and antigen-bearing dendritic cells (DCs) [
32], which control their migration into SLOs towards the gradient of CXCL13 [
26,
33,
34,
35]. CXCR5 regulates Burkitt’s lymphoma (BL) lymphomagenesis, B cell differentiation, and migration [
25,
29,
34].
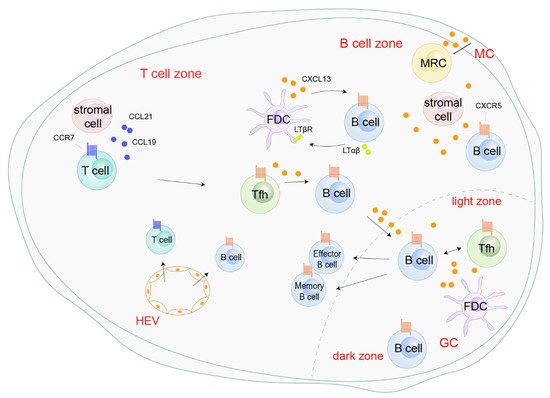
In response to CXCL13 secreted by Tfh, FDCs, or marginal reticular cells (MRC), and peripheral CXCR5+ B cells are recruited into the lymphoid follicles or germinal center (GC) in the SLOs through high endothelial venules (HEVs) (Figure 2). In the lymphoid follicles, a positive feedback loop mediated by CXCL13 boosts follicle development and sustains SLOs homeostasis. On one hand, CXCL13 secreted by FDCs upregulates membrane lymphotoxin α1β2 (LTαβ) on B cells. On the other hand, LTαβ interacts with the lymphotoxin-β receptor (LTβR) on FDCs and triggers FDC development, mutation, and CXCL13 production (Figure 2).
Figure 2. Roles of the CXCL13/CXCR5 axis in secondary lymphoid organs. CXCL13, secreted by follicular helper T cells (Tfh), follicular dendritic cells (FDCs), and marginal reticular cells (MRC), recruits peripheral CXCR5
+ B cells into the B cell zone or germinal center (GC) through high endothelial venules (HEVs). In the B cell zone, CXCL13 enhances follicle development and sustains secondary lymphoid organ homeostasis by a positive-feedback loop with B cells and FDC [
26]. CXCR5
+ Tfh migrate into B cell zones, initiating GC formation and B cell receptor (BCR) affinity maturation and promoting the differentiation of B cells into antibody-producing plasma cells and memory cells [
40,
41,
42].
3. CXCL13/CXCR5 and Non-Cancerous Diseases
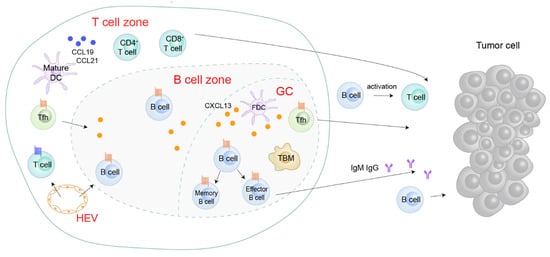
CXCL13 signaling is involved in multiple diseases and exhibits context-dependent effects in inflammatory conditions and tumor tissues. Generally, the tertiary lymphoid structure (TLS) will develop in non-lymphoid tissues within or near the pathological sites when an organism suffers from disorders, including persistent infection, autoimmune disease, chronic obstructive pulmonary disease (COPD), and cancer (
Figure 3) [
45,
46]. Structurally, TLSs have B cell zones, T cell zones, GCs, and HEVs.
Figure 3. Roles of the CXCL13/CXCR5 axis in the tertiary lymphoid structure (TLS). CXCL13 is aberrantly expressed in the TLS. CXCL13 attracts CXCR5
+ Tfh and B cells to the B cell zone or GC, potentiating B cell maturation and TLS formation [
47,
48]. In TLS, B cells’ secret immunoglobulins activate T cells or directly target cancer cells [
15,
49]. Tfh, follicular helper T cells; FDC, follicular dendritic cells; GC, germinal center; HEV, high endothelial venules; TBM, tingible body macrophages.
CXCL13 is aberrantly expressed and acts as the main orchestrator in TLSs [
24,
45,
47,
50,
51,
52,
53,
54]. After viral infection, type I interferon is produced and can induce CXCL13 production in a population of lung fibroblasts, driving CXCR5-dependent recruitment of B cells and initiating ectopic germinal center formation [
55]. Primary pulmonary fibroblast-secreted CXCL13 induces the formation of inducible bronchus-associated lymphoid tissue (iBALT), which drives immune responses to fungal stimulation within the lungs [
47]. In one fatal and irreversible interstitial lung disease, idiopathic pulmonary fibrosis, CXCL13 is produced by CD68- and CD206-positive alveolar macrophages, and the serum CXCL13 concentration predicts the progression and severity of the disease [
56]. In the lungs of mice and patients bearing COPD induced by chronic cigarette smoke exposure, CXCL13 is elevated in the lymphoid follicles and mediates the formation of TLS, resulting in chronic inflammation in bronchoalveolar lavage and destruction of alveolar walls [
45].
In the synovial tissues of rheumatoid arthritis, CXCL13 is produced by PD-1
hiCXCR5
−CD4
+ T cells [
57]. CXCL13 is also a key regulator of B cell recruitment to the cerebrospinal fluid in acute Lyme neuroborreliosis [
58]. CXCL13 participates in TLS formation in some autoimmune diseases, such as primary Sjögren’s syndrome [
59,
60], systemic lupus erythematosus, myasthenia gravis [
61], and atherosclerosis [
62], but not in rheumatoid arthritis [
63] or acute Lyme neuroborreliosis [
64]. CXCL13 is also highly produced during adipogenesis, and has been shown to be a differentiation- and hypoxia-induced adipocytokine that exacerbates the inflammatory phenotype of adipocytes through the induction of the pleckstrin homology (PH) domain leucine-rich repeat protein Ser/Thr specific phosphatase family 1 (PHLPP1), which regulates AKT activation [
65,
66].
CXCL13 is upregulated by
Helicobacter suis, the most prevalent non-
Helicobacter pylori species colonizing the stomach of humans suffering from gastric disease [
67]. CXCL13 is also induced in
Helicobacter-related chronic gastritis and is involved in the formation of lymphoid follicles and the gastric lymphomas of mucosa-associated lymphoid tissue types [
68,
69]. In non-alcoholic fatty liver disease (NAFLD), repressed expression of CXCL13 may ameliorate steatosis-related inflammation [
70]. CXCL13 drives spinal astrocyte activation and neuropathic pain via CXCR5 [
71], and is critical to preserve motor neurons in amyotrophic lateral sclerosis [
72]. Further understanding of the regulations and functions of the CXCL13/CXCR5 axis will aid the rational design of therapeutics for these diseases.
4. CXCL13/CXCR5 and Cancer
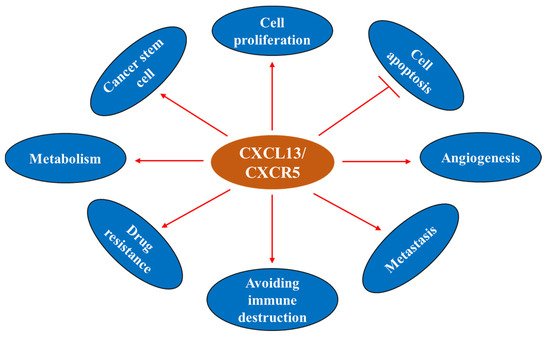
The CXCL13/CXCR5 axis is involved in the regulation of cancer cell survival, apoptosis, proliferation, differentiation, migration, invasion, and adaptive immunity, and shows dichotomic anti- and pro-tumor functions in the TME [
21,
50,
73,
74,
75,
76,
77,
78] (
Figure 4). CXCL13 and CXCR5 have important roles in cancer (
Figure 5) and represent potential markers to predict the response to immune checkpoint therapy [
51,
79,
80,
81]. These molecules may also serve as novel targets for the development of preventive and/or therapeutic agents for cancer.
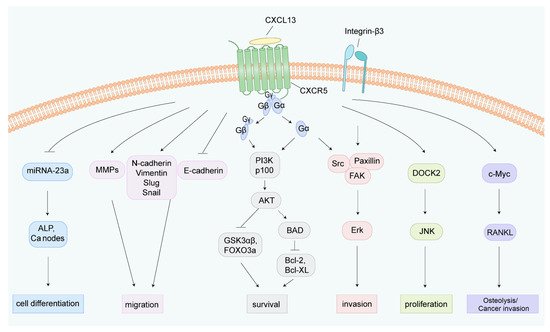
Figure 4. Illustration of the underlying mechanisms of the CXCL13/CXCR5 axis in cell fate determination. The CXCL13/CXCR5 axis triggers multiple intracellular signal transduction pathways. After CXCL13 binds to CXCR5, G proteins dissociate from CXCR5, dividing into Gα and Gβγ, thereby inducing different downstream molecular events [
20,
23]. CXCL13 promotes osteogenic differentiation by inhibiting miRNA-23a, inducing ALP activity, and calcium node formation [
82]. Upregulation of MMPs, N-cadherin, Vimentin, Slug, and Snail, and downregulation of E-cadherin under CXCL13 treatment enhances tumor cell migration [
18,
83]. The CXCL13/CXCR5 axis activates PI3K/Akt, integrin-β3/Src/Paxillin/FAK, and the DOCK/JNK pathway to induce cell survival, invasion, and proliferation, respectively [
73]. CXCL13 increases the phosphorylation of c-Myc and c-Jun, and upregulates the transcriptional regulator NFATc3, which binds to the promoter region of RANKL and elevates the expression of RANKL [
74,
84]. ALP, alkaline phosphatase; Ca, calcium; MMPs: matrix metalloproteinase; BAD, Bcl-2 agonist of cell death; FAK, focal adhesion kinase; DOCK2, dedicator of cytokinesis 2; JNK, c-Jun kinase; RANKL, receptor activator of NF-kB ligand.
Figure 5. CXCL13 and cancer hallmarks.
4.1. CXCL13 Sources within the Tumor and the Tumor Microenvironment
4.1.1. CXCL13: Cellular Sources within TME
CXCL13 is secreted by multiple populations of cells within the TME, including stromal cells, endothelial cells, lymphocytes, and tumor cells. FDC, a considerable stromal cell population, is the major producer of CXCL13 in the GCs [
85,
86]. Cancer-associated fibroblasts can convert to myofibroblasts and secrete CXCL13 into the TME upon hypoxia and TGF-β stimulation [
87]. CXCL13 is also produced by human bone marrow endothelial (HBME) cells [
88], Tfh that have infiltrated into tumor tissues [
89], PD1
+ CD8
+ T cells [
90], the TGFβ-dependent CD103
+CD8
+ tumor-infiltrating T-cell (TIL) subpopulation [
91], neoplastic T cells [
85], and several types of tumor cells.
4.1.2. CXCL13: Production under Carcinogen Stimulation
Environmental carcinogens can induce the production of CXCL13. Studies showed that the expression of CXCL13 at both the mRNA and protein levels was increased in B cell areas of lymphoid follicles in the lungs of cigarette smoke (CS)-exposed mice, and the CS-induced upregulation of CXCL13 was confirmed in patients with COPD. Interestingly, CS-induced formation of pulmonary lymphoid follicles was blocked by anti-CXCL13 antibodies in mice, and the absence of tertiary lymphoid organs (TLOs) in bronchoalveolar lavage alleviated the inflammatory response and destruction of the alveolar walls but did not impact the remodeling of the airway wall [
45].
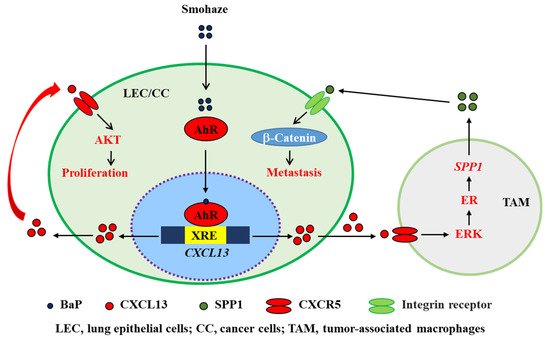
CXCL13 plays a critical role in environmental carcinogenesis. Wang et al. [
18] screened for abnormal inflammatory factors in patients with non-small cell lung cancers (NSCLCs) from Xuanwei city in China’s Yunnan Province, where the wide use of smoky coal resulted in severe household air pollution, and found that CXCL13 was substantially upregulated in 63 (90%) of 70 Yuanwei patients with NSCLC. In NSCLC patients from control regions where smoky coal was not used, CXCL13 was overexpressed in 44/71 (62%) of smoker patients and 27/60 (45%) of non-smoker patients [
18]. Benzo(a)pyrene (BaP), a polycyclic aromatic hydrocarbon (PAH) carcinogen found in tobacco smoking and haze (smohaze) [
92], can be metabolically activated by the production of BaP-7,8-diol-9,10-epoxides (BPDEs). BPDE reacts with DNA to form adducts at
N2 of deoxyguanosine (BPDE-
N2-deoxyguanosine), which induces mainly G→T genomic mutations to promote carcinogenesis [
93]. We found that BaP induced the production of CXCL13 by lung epithelial cells in vitro and in vivo. Consistent with these observations, CXCL13 was shown to be elevated in serum samples of current and former smokers and was associated with lung cancer risk [
94]. CXCL13 induces the production of secreted phosphoprotein 1 (SPP1 or osteopontin) by macrophages to activate β-catenin and induce an epithelial-to-mesenchymal transition (EMT) phenotype (
Figure 6). Deficiency in CXCL13 or CXCR5 significantly suppressed BaP-induced lung cancer in mice, indicating that CXCL13 plays a key role in smohaze carcinogen-induced lung cancers [
18,
92]. CXCL13 is also upregulated in human colorectal cancer and is secreted by dendritic cells [
95]. The carcinogen azoxymethane, which is catalyzed into methylazoxymethanol to induce G→A genomic mutations, induces colorectal cancer in vivo. Interestingly, knockout of CXCL13 inhibits azoxymethane/dextran sodium sulfate-induced colorectal cancer in mice [
95]. These data suggest a crucial role for the CXCL13-CXCR5 axis in cancers induced by environmental factors and could be a novel target for the development of preventive and therapeutic agents to combat related cancers.
Figure 6. Schematic representation of smohaze-induced production of CXCL13 in lung cancer.
4.2. CXCL13/CXCR5 and Cancer Hallmarks
4.2.1. CXCL13 and Cell Proliferation
CXCL13 binds specifically to CXCR5, which couples with MEK/ERK to induce cell proliferation [
96]. In clear cell renal cell carcinoma (ccRCC) cells, CXCL13 promotes proliferation by binding to CXCR5 and subsequently activating the PI3K/AKT/mTOR signaling pathway [
97]. The PI3K/AKT pathway also plays a key role in the CXCL13/CXCR5 axis, promoting colon cancer growth and invasion [
98,
99]. The CXCL13/CXCR5 axis promotes the proliferation and invasion of prostate cancer (PCa) cells by activating JNK, ERK, SRC/FAK, PI3K, and Akt [
73,
96,
100]. CXCL13 also promotes the proliferation of androgen-responsive LNCaP PCa cells in a JNK-dependent, DOCK2-independent manner, whereas in androgen-independent PC3 cells, CXCL13-induced proliferation is dependent on DOCK2 [
73,
100,
101].
4.2.2. CXCL13 and Cell Apoptosis
The CXCL13/CXCR5 axis plays an important role in cell homeostasis, as well as helping leukemic cells escape apoptosis by regulating chemokine-induced signaling [
103]. In breast cancer cells, the decrease in CXCL13 leads to the decreased expression of CXCR5, p-ERK/ERK, and cyclin D1 as well as the increased expression of cleaved Casp-9, which is an initiator caspase protease for apoptosis [
104]. Moreover, CXCL13/CXCR5 has been demonstrated to induce significant resistance to TNF-α-mediated apoptosis in B cell lineage acute and chronic lymphocytic leukemia (B-ALL and B-CLL) [
105,
106]. CXCL13 regulates the phosphorylation of Bcl-2 (at Serine 70), Bcl-xL (at Serine 62), and BAD (at Serine 112 and 136) in PC3 cells to exert anti-apoptotic effects [
73]. CXCR5 may be involved in the protection of retinal pigment epithelium (RPE) and retinal cells from aging-related photoreceptor apoptosis [
107]. These data demonstrate that the CXCL13/CXCR5 axis can confer the evasion of apoptosis in cancer cells by modulating p-ERK/ERK, TNF-α, Casp-9, and other signal pathways.
4.2.3. CXCL13 and Cancer Stem Cell (CSC)
Cancer stem cell (CSC) is a type of tumor cell with the abilities of self-renewal, differentiation, and high drug resistance [
108]. IL30 overproduction by prostate cancer stem cell-like cells promotes tumor initiation and development, which involves increased proliferation, vascularization, and myeloid cell recruitment. Moreover, it promotes stem cell-like cell dissemination to lymph nodes and bone marrow by upregulating the CXCR5/CXCL13 axis [
109]. CXCL13 recruits B cells to prostate tumors to promote castrate-resistant cancer progression by producing lymphotoxin, which activates an IκB kinase α (IKKα)-BMI1 module in prostate cancer stem cells [
110]. The role of the CXCL13/CXCR5 pathway in the cancer stem cells of other malignancies remains to be investigated.
4.2.4. CXCL13 and Drug Resistance
CXCL13/CXCR5 plays an essential role in drug resistance. In multiple myeloma (MM), CXCL13 secreted by mesenchymal stem cells (MSCs) confers resistance to bortezomib to MM cells [
111]. In 5-fluorouracil (5-Fu)-resistant colorectal cancer patients, serum CXCL13 is elevated, and a high CXCL13 concentration is associated with a worse clinical outcome [
112]. CXCL13 is significantly increased in diffuse large B-cell lymphoma resistance to chemotherapy and is involved in tumor progression [
113]. CXCR5 is overexpressed in mantle cell lymphoma (MCL), where it mediates MCL-stromal cell adhesion and drug resistance. The drug resistance of MCL is associated with increased expression of B-cell activation factor (BAFF), which induces the expression of CXCL13 [
114].
4.2.5. CXCL13/CXCR5 in the Tumor Microenvironment
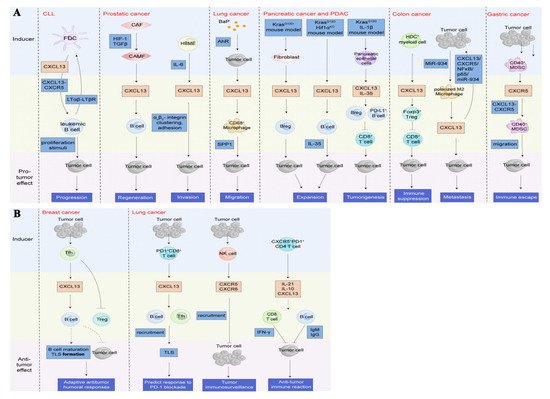
The CXCL13/CXCR5 axis may have different roles in the TME. In leukemia, prostate, lung, pancreatic, colon, and gastric cancers, CXCL13 exhibits pro-cancer effects by recruiting B cells [
86,
87,
115,
116], CD68
+ macrophages [
18], regulatory B cells (Bregs) [
117,
118], Treg [
119], and CD40
+ MDSCs [
120], shaping an immune-suppressive TME to trigger tumorigenesis and tumor progression (
Figure 7A). A few reports regarding breast and lung cancers have shown that the CXC13/CXCR5 axis attracts B cells and Tfh [
89,
90] to shape the TLS in the peritumoral or tumor sites (
Figure 7B), which is associated with adaptive anti-tumor humoral responses and predicting responses to PD-1 blockade therapy.
Figure 7. Roles of the CXCL13/CXCR5 axis in the tumor microenvironment (TME). (
A). The CXCL13/CXCR5 axis plays a crucial role in shaping a complex TME by recruiting multiple types of lymphocytes to beget pro-tumor or anti-tumor immunity reactions. In the context of pro-neoplastic reactions, the CXCL13/CXCR5 axis attracts B cells to induce tumor progression, regeneration, and invasion [
86,
87,
115,
118], or recruits CD68
+ macrophages [
18], Breg [
117,
118], Treg [
119], and CD40
+ MDSC [
120] to trigger migration, expansion and tumorigenesis, immune suppression, and immune escape, respectively. (
B). In the circumstance of anti-tumor reactions, the CXC13/CXCR5 axis recruits B cells and Tfh [
89,
90] to format TLS in the peritumoral or tumor sites, which is associated with adaptive anti-tumor humoral responses and predicting the response to PD-1 blockade therapy. Additionally, lymphocytes directly or indirectly dampen tumors by the upregulation of CXCL13 and/or CXCR5 [
15,
121]. CLL, chronic lymphocytic leukemia; LTαβ, lymphotoxin α1β2; LTβR, lymphotoxin-β receptor; CAF, cancer-associated fibroblasts; CAMF, cancer-associated myofibroblasts; HIF-1, hypoxia-inducible factor 1; TGFβ, transforming growth factor-β; HBME, human bone marrow endothelial; IL, interleukin; BaP, benzo(a)pyrene; AhR, Aryl hydrocarbon receptor; SPP1, secreted phosphoprotein 1; PDAC, pancreatic ductal adenocarcinoma; Breg, regulatory B cells; HDC, histidine decarboxylase; Treg, regulatory T cells; MDSC, myeloid-derived suppressor cells; Tfh, follicular helper T cells; NK cell, natural killer cell.
4.2.6. CXCL13 and Angiogenesis
Angiogenesis is a distinguishable characteristic of successful tumor growth in all solid tumors, and CXC chemokines are pleiotropic in their ability to regulate tumor-associated angiogenesis, as well as cancer cell metastases [
125]. Chronic hypoxia increases the expression of CXCL13 in adipocytes [
65] and promotes the metastasis of prostate cancer by increasing the expression of CXCL13 in tumor myofibroblasts [
87]. Fibroblast growth factor-2 (FGF2) is a member of the family of the heparin-binding FGF growth factors with pro-angiogenic activity. CXCL13 inhibits FGF2-induced chemotaxis and proliferation, as well as the survival of endothelial cells, acting as an angiostatic chemokine [
126]. CXCL13/CXCR5 axis also facilitates angiogenesis during rheumatoid arthritis progression [
127].
4.2.7. CXCL13 and Immunometabolic Responses
An integrated immunometabolic response during negative energy balance is required for host survival, and the impacts of nutritional status on immune responses remain to be determined. Recent studies have shown that temporary fasting significantly reduces the number of lymphocytes in Peyer’s patches, whose cellular composition is conspicuously altered after resuming feeding, with the numbers seemingly restored. In this process, nutritional signals are necessary to maintain CXCL13 expression by stromal cells [
128]. Fasting reduces the numbers of circulating monocytes, as well as monocyte metabolic and inflammatory activity, while hepatic energy-sensing regulates homeostatic monocyte numbers via CCL2 production [
129]. However, the potential roles CXCL13 plays in cancer metabolism remain to be investigated.
4.2.8. CXCL13 and Cancer Metastasis
More than 90% of cancer deaths are attributed to metastasis. The intricate interactions of a chemokine and its receptor play an essential role in tumor metastasis. CXCL13/CXCR5 also participates in the metastasis of multiple cancers. CXCL13 enhances cancer metastasis signaling in an autocrine or paracrine manner, since it is secreted by tumor cells or other cell types, such as stromal cells and lymphocytes. In a murine prostate cancer model, which exhibits PKCε overexpression and
Pten deficiency, the release of CXCL13 by tumor cells was upregulated in a non-canonical NF-κB pathway, boosting tumor cells’ migratory properties [
122]. CXCL13 facilitates breast cancer cell line migratory activity via the nuclear factor kappa-B ligand (RANKL)-Src pathway, which mediates the upregulation of EMT regulators and matrix metalloproteinase-9 (MMP9) [
83,
130]. CXCL13, also secreted by stromal cells, upregulates the expression of RANKL on stromal cells, promoting tumor cell migration and lymph node metastasis via the RANK-RANKL pathway [
131]. CXCL13 mediates distal metastasis of colon cancer by increasing the secretion of MMP13 and the activation of the PI3K/Akt pathway [
132].
4.3. Regulation of CXCL13 in Tumors
A series of studies have shed new light on the regulation of CXCL13 and CXCR5 in tumors. RelA, a subunit of the NF-κB family [
134], directly binds to the CXCL13 promoter and positively regulates the transcription of CXCL13, while nuclear factor erythroid 2-related factor 2 (NRF2) acts as a negative transcriptional regulator of this chemokine [
135]. CXCR5 was positively regulated by RelA and negatively by p53 [
135,
136], and nuclear raf-1 kinase regulates the CXCR5 promoter by associating with NFATc3 [
137]. P53 homologues, p63 and p73 [
138], utilize the same mechanism by which the activity of NFκB is attenuated to reduce the expression of CXCR5 [
139]. The aryl hydrocarbon receptor (AhR), a ligand-activated transcription factor, is translocated to the nucleus under BaP stimulation and binds to the xenobiotic-responsive element (XRE) in the promoter of
CXCL13, positively regulating the transcription of CXCL13 (
Figure 6) [
18].
5. CXCL13/CXCR5 in Several Cancer Types
Mounting evidence demonstrates a high concentration of CXCL13 and/or high expression of CXCR5 in tumor tissues or tumor cell lines. The CXCL13/CXCR5 axis in both hematological malignancies and solid tumors mediates multiple intracellular signal cascade reactions and yields various phenotypes responding to the signaling pathways. In addition, the CXCL13/CXCR5 axis also potentiates the crosstalk between tumor cells and lymphocytes or non-lymphocytes, shaping a complex TME. The roles of the CXCL13/CXCR5 axis participating in the malignant tumors are context-dependent, including pro-tumor and anti-tumor activities (
Figure 7). On one hand, CXCL13 attracts immunosuppressive cells to mediate immune suppression or evasion, leading to tumor progression, while on the other hand, the CXCL13/CXCR5 axis elicits tumoricidal immunity signaling to escape tumor immunosurveillance in some cancer types [
54,
89,
141] (
Table 1).
Table 1. Therapeutic targets associated with the CXCL13/CXCR5 axis in malignancies and the tumor microenvironment.
|
Target
|
Cancer Type
|
Function
|
Approach
|
In Vivo or In Vitro
|
Outcome
|
Refs.
|
|
CXCL13
|
Prostate cancer
|
Induction of prostate cancer cell proliferation and migration
|
siRNA and shRNA; antibody
|
In vivo; in vitro
|
Inhibiting tumor growth and metastasis
|
[122]
|
|
CXCL13
|
Prostate cancer
|
Chemotaxis B cells into regressing tumor
|
Antibody
|
In vivo
|
Preventing B-cell recruitment into tumor under castration
|
[115]
|
|
CXCL13
|
Breast cancer
|
Activating CXCR5/ERK pathway
|
Polyclonal antibody
|
In vivo; in vitro
|
Attenuating tumor volume and growth; inhibiting tumor cell proliferation and promoting its apoptosis
|
[102,104]
|
|
CXCL13
|
Breast cancer
|
Enhancing the production of RANKL on tumor cells and the interaction between ILC3 and stromal cells
|
Antibody
|
In vivo
|
Attenuating lymph node metastasis
|
[131]
|
|
CXCL13
|
Lung cancer
|
Promotion of cell proliferation; inducing the production of SPP1 by microphage
|
Cxcl13−/− mice
|
In vivo
|
Decreasing the volume of BaP-induced tumor
|
[18]
|
|
CXCL13
|
PDAC
|
Homing B cell into tumor lesions
|
Antibody
|
Mice harbored KrasG12D PDEC
|
Reducing the growth of orthotopic tumor
|
[116]
|
|
CXCL13
|
Colon cancer
|
Induction 5-Fu resistance and association with a worse outcome
|
siRNA
|
In vitro
|
Reducing 5-Fu resistance
|
[112]
|
|
CXCR5
|
CLL
|
CXCR5+ leukemia B cells recruited by CXCL13 to encounter proliferation stimuli
|
Cxcr5−/− Eμ-Tcl1 mice
|
In vivo
|
Attenuating tumor cell proliferation
|
[86]
|
|
CXCR5
|
Prostate cancer
|
Induction of prostate cancer cells proliferation and migration
|
siRNA and shRNA
|
In vivo; in vitro
|
Inhibiting tumor growth and metastasis
|
[122]
|
|
CXCR5
|
Lung cancer
|
CXCR5+ CD68+ macrophages producing SPP1 to promote EMT process
|
Cxcr5−/− mice
|
In vivo
|
Decreasing the volume of BaP-induced tumor
|
[18]
|
|
CXCR5
|
OSCC
|
Induction RANKL expression under CXCL13/CXCR5 axis
|
Antibody
|
In vitro
|
Inhibiting the expression of RANKL
|
[74]
|
|
TGFβR
|
Prostate cancer
|
Activating CXCL13-expressing myofibroblasts
|
SB-431542
|
In vivo
|
Blocking the initiation of castration-resistant prostate cancer
|
[87]
|
|
NFATc3
|
OSCC
|
Nuclear translocation mediated by CXCL13/CXCR5 axis to bind to RANKL promoter region
|
siRNA
|
In vitro
|
Preventing RANKL expression
|
[74]
|
|
Myofibroblasts
|
Prostate cancer
|
Induction of CXCL13 expression
|
Immunodepletion; phosphodiesterase 5
|
In vivo
|
Blocking the initiation of castration-resistant prostate cancer
|
[87]
|
ILC3, RORγt+ innate lymphoid cell group 3; SPP1, secreted phosphoprotein 1; BaP, benzo(a)pyrene; PDAC, pancreatic ductal adenocarcinoma; PDEC, pancreatic ductal epithelial cells; 5-Fu, 5-Fluorouracil; CLL, chronic lymphocytic leukemia; EMT, epithelial to mesenchymal transition; OSCC, oral squamous cell carcinomas; RANKL, RANK ligand; TGFβR, TGFβ receptor; NFATc3, nuclear factor of activated T cells.
This entry is adapted from the peer-reviewed paper 10.3390/life11121282