During the past two decades, there have been significant advances in nucleic acid carriers modified by histidines or histidine-rich domains. There are several properties of histidines, primarily emanating from their buffering of acidic endosomes, which augment transfection. These roles from protonated histidines include osmotic swelling with lysis of endosomes, unpacking of the carrier complex, and release of the nucleic acids to enable the carrier to interact with the endosomal membrane. Histidines or histidine-rich peptides have been incorporated into polymers, conjugated to lipids, phages, and mesoporous silica particles, as well as formed shields around nanoparticles. These carriers have demonstrated significant potential to import into the cytosol different forms of nucleic acids including plasmids, siRNA, and mRNA.
- peptide
- histidine
- imidazole
- nucleic acids
- siRNA
- plasmid
- mRNA
- \
1. Introduction
Histidines incorporated into carriers of nucleic acids may enhance the extracellular stability of the nanoparticle, yet aid in the intracellular disruption of the nanoparticle, enabling the release of the nucleic acid. Moreover, protonation of histidines in the endosomes may result in endosomal swelling with subsequent lysis. These properties of histidine are based on its five-member imidazole ring in which the two nitrogen atoms may form hydrogen bonds or act as a base in acidic environments. A wide variety of carriers have integrated histidines or histidine-rich domains, which include peptides, polyethylenimine, polysaccharides, platform delivery systems, viral phages, mesoporous silica particles, and liposomes. In the review [1], we have divided histidine-rich carriers of nucleic acids into several categories (see below). These histidine-rich carriers or their polyplex cores may be further modified by PEGylation, cross-linkers including oxidized cysteines, or targeting ligands to enhance transfection.
2. Histidine-Lysine Carriers of Nucleic Acids
With the single repeating subunit-containing both DNA-condensing and pH buffering properties, polyethylenimine and polyamidoamine are efficient nucleic acid delivery agents. Except for altering the degree of branching, the binding and buffering properties of the two polymers cannot be readily varied. Consequently, there may be an inherent inflexibility of these polymers, which have a single repeating subunit with two or more functional features. In contrast, the ratios of histidines/imidazoles and lysines can be readily modified, affecting the ability of these histidine-lysine carriers to transport different forms of nucleic acids. Whereas cationic lysines bind to the negatively charged phosphates of nucleic acids, histidines and imidazoles have a number of roles as delineated in the introduction. Several studies have demonstrated the efficacy of nucleic acid carriers consisting primarily of histidines and lysines [2-6]. The seminal study of Midoux and Mosigny launched the field in which they incorporated the histidine moiety into a polylysine transfection carrier (Figure 1a) to enhance gene expression in cell culture [2]. Moreover, our lab demonstrated that specific sequences of histidines and lysines were important in the transport of nucleic acids [4,5] (Figure 1b).
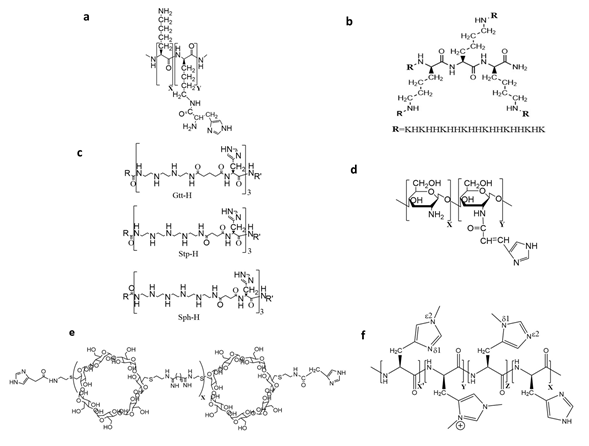
Figure 1. Chemical structures of selective histidine-rich polymers. (a) Histidylated polylysine, (b) four-branched histidine-lysine peptide with a 3-lysine core, (c) protonated and buffering subunits (Gtt-H, Stp-H, and Sph-H) incorporated into linear and branched polymers, (d) urocanic-modified chitosan, (e) cyclodextrin-containing polycation modified by two imidazole groups, (f) the imidazole ring of polyhistidine modified by methylation. The number of unmodified (X) and modified (Y, Z, X’) monomeric units in the polymer (a,d–f) are indicated.
3. Beyond H-K Peptides and Polymers
In addition to lysines, other amino acids and modifications have been incorporated into histidine-rich peptides and polymers to improve these carriers of nucleic acids. Amino acids such as leucine, alanine, ornithine, and arginine, as well as non-amino modifications including diethylamines or aminoethyl modified histidines, have been used [7-12]. For example, Lachelt et al. examined several linear and branched cationic oligoethanamino amide as carriers of plasmids (Figure 1c) [10]. Instead of lysines that bind to nucleic acids, the cationic segments of these oligoethanamino amide polymers were made up of diaminoethanes, the same repeating motif that is in PEI.
There are also several examples of histidine/imidazoles or histidine-rich peptides, which have been added to parent carriers such as PEI, PAMAM, TAT, chitosan (Figure 1d) and cyclodextrin (Figure 1e), to enhance the delivery of nucleic acids [13-17]. Histidine/imidazole-containing peptides have been added to the parent non-biodegradable and biodegradable carriers for several reasons. For non-biodegradable carriers such as PEI, an unheralded role of histidines is its ability to reduce the toxicity of polymers and polyplexes [13–15]. By conjugating imidazoles to branched PEI, reduced toxicity of the modified PEI was observed in vivo and was mediated by a decrease of cytokines, chemokines, and reduced liver injury [15].
4. Modification of Polyhistidines
When the pH is above 6.0, the solubility of polyhistidine is greatly reduced and the stability of such polyplexes remains a challenge. Thus far, several strategies have been discussed that have incorporated lysines or other cationic agents with histidines/imidazoles in carriers to bind DNA and increase the solubility of the carrier and the polyplex. Alternatively, there have been approaches to enhance the solubility and stability of polyhistidine polyplexes, particularly when these are formed at neutral pH. These modifications of polyhistidine include PEGylation, glycosylation, amination, methylation (Figure 1f), and most recently, by carboxymethyl groups [18].
5. Lipopeptides
Lipopeptide or lipid-peptide hybrid carriers form micelles alone and in complex with nucleic acids. Since the earliest report of lipid-peptide hybrids, these hybrids have evolved into an important area for histidine-rich polymeric carriers. For instance, Porosk et al determined that some histidine patterns that were N-terminal to the KALA group were effective carriers in vitro and in vivo. After a single intravenous injection, one histidine-containing analog silenced factor VII in the liver of mice by about 60% compared to untreated controls [19]. Alternatively, Li and colleagues co-delivered in histidine-rich micelles a siRNA targeting PD-1 and a small molecule inhibitor (methyl-tryptophan) to reduce tumor size [20].
6. Lipopolyplexes/Lipoplexes
Histidine-rich peptides combined with cationic liposomes (lipopolyplexes or lipoplexes) are closely related to histidine-rich peptide-lipid hybrids. Several studies have demonstrated the efficacy of using these histidine-rich carriers. Of particular interest is their promise as carriers of mRNA vaccines targeting cancer. In these reports, decorating the liposome surface with a “tri-antenna of α-D-mannopyranoside” (Tri-man) improved the specificity of the mRNA vaccine toward dendritic cells compared to those decorated with mannose and markedly improved the antitumor response. This platform has significant promise as a delivery system for therapeutic cancer vaccines in humans [21].
7. Pre-Formed Nanoparticles and Virus-Like Particles Modified with Imidazoles
In addition to liposomes, non-liposomal preformed vectors have been modified with imidazoles/histidines or histidine-rich peptides as carriers of nucleic acids. These preformed nanoparticles include mesoporous silica particles (MSN) and viral-like particles. In one report, imidazoles were conjugated via a Schiff base linkage to mesoporous silica nanoparticles to enhance the delivery of survivin shRNA-expressing plasmids and doxorubicin to tumors. The imidazole MSN nanoparticles co-delivering the survivin-shRNA and doxorubicin completely inhibited the growth of the tumors [22]. Alternatively, an imidazole-zinc framework of a few nanometers of thickness, which covered the surface of the mesoporous silica nanoparticles, enabled the efficient loading of siRNA [23].
8. Conclusion
Over the past 20 years, there have been significant advances in nucleic acid carriers modified by histidines or histidine-rich domains. There are several properties of histidines, primarily emanating from their buffering of acidic endosomes, which increase transfection. Thus far, histidines or histidine-rich peptides have been incorporated into polymers, conjugated to lipids, phages, and mesoporous silica nanoparticles, as well as formed shields around nanoparticles. Although the addition of histidines to diverse nanoparticles has markedly improved their transfection, understanding the interactions of histidine-rich domains with nucleic acids and biological membranes will likely be necessary to realize the full potential of histidine-modified nanoparticles.
References
- He, J.; Xu, S.; Mixson, A.J. The Multifaceted Histidine-Based Carriers for Nucleic Acid Delivery: Advances and Challenges. Pharmaceutics 2020, 12, doi:10.3390/pharmaceutics12080774.
- Midoux, P.; Monsigny, M. Efficient gene transfer by histidylated polylysine/pDNA complexes. Chem. 1999, 10, 406–411, doi:10.1021/bc9801070.
- Putnam, D.; Gentry, C.A.; Pack, D.W.; Langer, R. Polymer-based gene delivery with low cytotoxicity by a unique balance of side-chain termini. Natl. Acad. Sci. USA 2001, 98, 1200–1205, doi:10.1073/pnas.031577698.
- Chen, Q.R.; Zhang, L.; Luther, P.W.; Mixson, A.J. Optimal transfection with the HK polymer depends on its degree of branching and the pH of endocytic vesicles. Nucleic Acids Res. 2002, 30, 1338–1345, doi:10.1093/nar/30.6.1338.
- Leng, Q.; Mixson, A.J. Modified branched peptides with a histidine-rich tail enhance in vitro gene transfection. Nucleic Acids Res. 2005, 33, e40, doi:10.1093/nar/gni040.
- Canine, B.F.; Wang, Y.; Hatefi, A. Evaluation of the effect of vector architecture on DNA condensation and gene transfer efficiency. Control. Release 2008, 129, 117–123, doi:10.1016/j.jconrel.2008.04.012.
- Kichler, A.; Leborgne, C.; Marz, J.; Danos, O.; Bechinger, B. Histidine-rich amphipathic peptide antibiotics promote efficient delivery of DNA into mammalian cells. Natl. Acad. Sci. USA 2003, 100, 1564–1568, doi:10.1073/pnas.0337677100.
- Chamarthy, S.P.; Kovacs, J.R.; McClelland, E.; Gattens, D.; Meng, W.S. A cationic peptide consists of ornithine and histidine repeats augments gene transfer in dendritic cells. Immunol. 2003, 40, 483–490.
- Lo, S.L.; Wang, S. An endosomolytic Tat peptide produced by incorporation of histidine and cysteine residues as a nonviral vector for DNA transfection. Biomaterials 2008, 29, 2408–2414, doi:10.1016/j.biomaterials.2008.01.031.
- Lachelt, U.; Kos, P.; Mickler, F.M.; Herrmann, A.; Salcher, E.E.; Rodl, W.; Badgujar, N.; Brauchle, C.; Wagner, E. Fine-tuning of proton sponges by precise diaminoethanes and histidines in pDNA polyplexes. Nanomedicine 2014, 10, 35–44, doi:10.1016/j.nano.2013.07.008.
- Zhang, T.T.; Kang, T.H.; Ma, B.; Xu, Y.; Hung, C.F.; Wu, T.C. LAH4 enhances CD8+ T cell immunity of protein/peptide-based vaccines. Vaccine 2012, 30, 784–793, doi:10.1016/j.vaccine.2011.11.056.
- Jia, L.; Kovacs, J.R.; Zheng, Y.; Gawalt, E.S.; Shen, H.; Meng, W.S. Attenuated alloreactivity of dendritic cells engineered with surface-modified microspheres carrying a plasmid encoding interleukin-10. Biomaterials 2006, 27, 2076–2082, doi:10.1016/j.biomaterials.2005.09.032.
- Gomez, J.P.; Tresset, G.; Pichon, C.; Midoux, P. Improved histidinylated lPEI polyplexes for skeletal muscle cells transfection. J. Pharm. 2019, 559, 58–67, doi:10.1016/j.ijpharm.2019.01.003.
- Salmasi, Z.; Shier, W.T.; Hashemi, M.; Mahdipour, E.; Parhiz, H.; Abnous, K.; Ramezani, M. Heterocyclic amine-modified polyethylenimine as gene carriers for transfection of mammalian cells. J. Pharm. Biopharm. 2015, 96, 76–88, doi:10.1016/j.ejpb.2015.07.008.
- Toy, R.; Pradhan, P.; Ramesh, V.; Di Paolo, N.C.; Lash, B.; Liu, J.; Blanchard, E.L.; Pinelli, C.J.; Santangelo, P.J.; Shayakhmetov, D.M.; et al. Modification of primary amines to higher order amines reduces in vivo hematological and immunotoxicity of cationic nanocarriers through TLR4 and complement pathways. Biomaterials 2019, 225, 119512, doi:10.1016/j.biomaterials.2019.119512.
- Kim, T.H.; Ihm, J.E.; Choi, Y.J.; Nah, J.W.; Cho, C.S. Efficient gene delivery by urocanic acid-modified chitosan. Control. Release 2003, 93, 389–402, doi:10.1016/j.jconrel.2003.08.017.
- Mishra, S.; Heidel, J.D.; Webster, P.; Davis, M.E. Imidazole groups on a linear, cyclodextrin-containing polycation produce enhanced gene delivery via multiple processes. Control. Release 2006, 116, 179–191, doi:10.1016/j.jconrel.2006.06.018.
- Gu, J.; Wang, X.; Jiang, X.; Chen, Y.; Chen, L.; Fang, X.; Sha, X. Self-assembled carboxymethyl poly (L-histidine) coated poly (beta-amino ester)/DNA complexes for gene transfection. Biomaterials 2012, 33, 644–658, doi:10.1016/j.biomaterials.2011.09.076.
- Porosk, L.; Arukuusk, P.; Pohako, K.; Kurrikoff, K.; Kiisholts, K.; Padari, K.; Pooga, M.; Langel, U. Enhancement of siRNA transfection by the optimization of fatty acid length and histidine content in the CPP. Sci. 2019, doi:10.1039/c9bm00688e.
- Li, G.; Gao, Y.; Gong, C.; Han, Z.; Qiang, L.; Tai, Z.; Tian, J.; Gao, S. Dual-Blockade Immune Checkpoint for Breast Cancer Treatment Based on a Tumor-Penetrating Peptide Assembling Nanoparticle. ACS Appl. Mater. Interfaces 2019, 11, 39513–39524, doi:10.1021/acsami.9b13354.
- Le Moignic, A.; Malard, V.; Benvegnu, T.; Lemiegre, L.; Berchel, M.; Jaffres, P.A.; Baillou, C.; Delost, M.; Macedo, R.; Rochefort, J.; et al. Preclinical evaluation of mRNA trimannosylated lipopolyplexes as therapeutic cancer vaccines targeting dendritic cells. Control. Release 2018, 278, 110–121, doi:10.1016/j.jconrel.2018.03.035.
- Li, Z.; Zhang, L.; Tang, C.; Yin, C. Co-Delivery of Doxorubicin and Survivin shRNA-Expressing Plasmid Via Microenvironment-Responsive Dendritic Mesoporous Silica Nanoparticles for Synergistic Cancer Therapy. Res. 2017, 34, 2829–2841, doi:10.1007/s11095-017-2264-6.
- Pan, Q.S.; Chen, T.T.; Nie, C.P.; Yi, J.T.; Liu, C.; Hu, Y.L.; Chu, X. In Situ Synthesis of Ultrathin ZIF-8 Film-Coated MSNs for Codelivering Bcl 2 siRNA and Doxorubicin to Enhance Chemotherapeutic Efficacy in Drug-Resistant Cancer Cells. ACS Appl. Mater. Interfaces 2018, 10, 33070–33077, doi:10.1021/acsami.8b13393.
This entry is adapted from the peer-reviewed paper 10.3390/pharmaceutics12080774