microRNAs (miRNAs) have been documented to be regulators of valvular diseases pathogenesis, diagnostic biomarkers, and therapeutical targets. They in fact play stimulatory or inhibitory roles in mitral valve prolapse development, aortic leaflet fusion, and calcification pathways. Tissue expression assessment and comparison between physiological and pathological phenotypes of different disease entities, including mitral valve prolapse and mitral chordae tendineae rupture, emerged as the best strategies to address miRNAs over or under-representation and thus, their impact on pathogeneses.
miRNAs can also be targeted by several molecules. Inhibitors such as antisense oligonucleotides and sponge vectors are under investigation. Furthermore, to increase miRNAs activity miRNA mimics, miRNA expression vectors, and small molecules have been developed.
1. miRNAs Effects on Aortic Stenosis and Mitral Prolapse Necessity of Novel Biomarkers
In aortic stenosis, miRNAs were also found to be altered in stenotic aortic valve leaflets compared to insufficient ones by Nigam et al. [6]. The reduced levels of miR-26a, miR-30b, and miR-195 reported in stenotic valves, were almost certainly responsible for their higher risk of developing valvular leaflet fusion. The change of morphology is due to accelerated calcium accumulation compared to regurgitant aortic valves without morphological fusion of valve leaflets. These miRNAs were involved in the biological processes that modulate calcification-related genes in vitro [6]. Other studies have confirmed miRNAs’ involvement in calcification pathways and will be discussed later.
Several studies have then recorded an important association of miRNAs with post-transcriptional regulation of gene expression in aortic valve stenosis. miR-141 was found to be a regulator of the levels of bone morphogenetic protein 2 (BMP-2), whereby unrestrained activity led to calcification of the aortic valve mediated by a stimulation of osteogenesis. miR-141 was markedly attenuated in patients with aortic stenosis associated with the bicuspid aortic valve [7]. Yanagawa et al. proposed this new key role of miR-141 in the modulation of aortic valve calcification disorders, highlighting the strategic therapeutic target that emerged in the assessment of progressive calcification in stenotic aortic valve disease [6,7]. The peculiar morphologic features of the stenotic aortic valve may probably be explained by the lower expression of miR-30b which is a known repressor of bone morphogenetic protein 2-mediated osteogenesis [7].
Zhang et al. demonstrated the role of miR-30b in reducing osteoblast differentiation activity induced by bone morphogenetic protein 2 [8]. The latter was implicated in promoting calcific aortic valve disease. The expression of miR-30b was found effective in reducing the risk of human aortic valve calcification and apoptosis through direct targeting of Runx2, Smad1, and caspase-3 [8] ( Figure 1 ).
Plasma levels of miRNAs have been used to monitor degenerative disease of the mitral valve (MV) but have not been widely adopted by the cardiology community [4,6,7,9,10,11]. The lack of enthusiasm of cardiologists for investigations into degenerative aortic valve stenosis [6,7,8,12,13] can be partially deduced from the lack of positive findings in mitral valve degenerative diseases leading to regurgitation [14,15,16,17,18,19].
2. Altered miRNAs Expression in Congenital Valve Disorders and Cardiogenetic Processes
Congenital heart diseases (CHD) comprise a large group of functional and structural disorders, namely atrial septal defects (ASD), ventricular septal defects (VSD), pulmonary valve atresia (PVA), coarctation of the aorta (CoA), tricuspid atresia (TA), tetralogy of Fallot (TOF) and several others [
63,
64,
65]. miRNAs play a pivotal role in heart development. The process of cardiac tissue formation and expression requires precise regulation and single miRNAs studies have been addressed in past decades. miR-1 and miR-133 are transcribed in a tissue-specific manner during development [
66]. MiR-1 targets HDAC4, histone deacetylase 4 which is a repressor for muscle gene expression, thereby stimulating myogenesis. miR-133 inhibits serum response factor (SRF) and promotes the differentiating process [
66].
Several other preclinical studies, conducted on zebrafish ventricles, denoted the role of miR-133 to diminish cardiac regenerating processes [
67]. Following resection of zebrafish ventricular apex, a reduced expression of miR-133, coupled with increased regenerative potential, has led to this concept [
67].
Cardiogenesis is also suppressed by the miR-15 family, which, specifically inhibited, has shown to promote myocyte proliferation after myocardial infarction [
68]. On the contrary, miR-199 and miR-590 have been found to promote the re-entry of cardiomyocytes in the cell cycle. An interesting therapeutical strategy, supported by preclinical mice studies, would be to inject these molecules into the border zones of infarcted hearts. Positive results and stimulation of cardiomyocytes proliferation have been demonstrated [
69,
70].
Several miRNAs also regulate the signals of insulin-growth factor 1 (IGF-1) in skeletal muscle, contributing to muscle development or atrophy [
71]. Several other studies have noted their regenerative role by observing common cardiovascular pathologies and the subsequent structural remodeling [
72]. Long non-coding RNAs are implied in hypertension-related vascular remodeling, post-ischemic recovery, and myocardial hypertrophy [
72].
Specific miRNAs are also differentially expressed in bicuspid aortic valve (BAV), the most common congenital heart disease. Aortic valve endothelial cells on the ventricular side are frequently exposed to high shear forces while on the aortic side turbulent blood flow and high levels of antioxidant enzymes are present [
73]. Conversely, on the ventricular side, factors inhibiting calcification are more abundant [
73,
74].
MiR-133 can be used for guiding the therapeutic management of aortic stenosis, due to its potential role in predicting left ventricular hypertrophy [
75,
76,
77]. Sabatino et al. performed a bioinformatic analysis to identify the most commonly regulated miRNAs in normal and stenotic bicuspid aortic valves and compared results with normal and stenotic tricuspid valves for calcium metabolism, blood coagulation, phosphate metabolism, and inflammatory pathways [
13]. The authors’ noted that the levels were differentially expressed in bicuspid versus tricuspid aortic valves and were also correlated with the degree of stenosis, as previously discussed. The investigators claim it will be used as a biomarker as it reflects the degree of myocardial fibrosis [
13].
The key factor involved in calcium metabolism and inflammatory pathways was found to be pidermal growth factor receptor (EGFR) [
78]. Several miRNAs, also associated with calcification, were associated with stenotic tricuspid aortic valves (TAVs) and BAVs, namely miR-100, -130a, -181a/181b, -199a-5p, -199a-3p, and -214 which have been investigated by other authors to display higher expression levels in VECs of the fibrosa on the aortic side, compared to the ventricular side [
79].
miR-181 is another important miRNA. Aortic valve endothelial cells were associated with its increased expression but decreased levels of targets, including SIRT1 and GATA6 that negatively affect vascular SMCs elastin production [
80]. Several other studies confirmed that its inhibition increases the expression of elastin and collagen while its stimulation, through direct administration, inhibits atherosclerotic lesion formation [
81].
3. Novel Therapeutical Strategies: miRNAs Targeting to Suppress or Activate Them
3.1. Results from In-Vivo and In-Vitro Testing for Aortic Valvular Stenosis
Regulating miRNAs expression is an attractive therapeutic challenge. Valvular calcification is currently not a direct target for pharmacological action. Endothelin receptor antagonists have emerged as possible molecules of interest in preclinical studies [
81,
82]. Chronic kidney disease-induced valvular calcification was demonstrated to be slowed by administration of endothelin type A (ETA) receptor atransentan (10 mg/kg/day) which reduced SMC differentiation, calcification, and stiffness [
81].
Concerning statin treatment, several studies reported increased rates of calcification [
83]. Possible strategies for miRNAs overexpression include miRNA mimics, miRNA expression vectors, and small molecules [
52,
84]. Instead, negatively regulating mi-RNAs seems to encompass different strategies. Antisense oligonucleotides including locked nucleic acid (LNA)-modified anti-miR, or miRNA sponge vectors can be used to specifically bind to miRNAs [
52]. Toshima et al. demonstrated miR-34a as a potential therapeutic target. Its inhibition in human aortic tissue exhibiting either calcific aortic valve stenosis (CAVS) or aortic regurgitation (AR) attenuated calcification signals in porcine aortic valve interstitial cells (AVICs) compared with miR-control [
85]. After performing RNA pull-down assays, miR-34a was demonstrated to directly target Notch1 by binding to Notch1 mRNA 3′ untranslated region [
85]. Additionally, miR-34a inhibitor suppressed calcium deposition of aortic valves and cardiac hypertrophy, both mechanisms involved in decreased Runx2 and increased Notch1 expressions [
85].
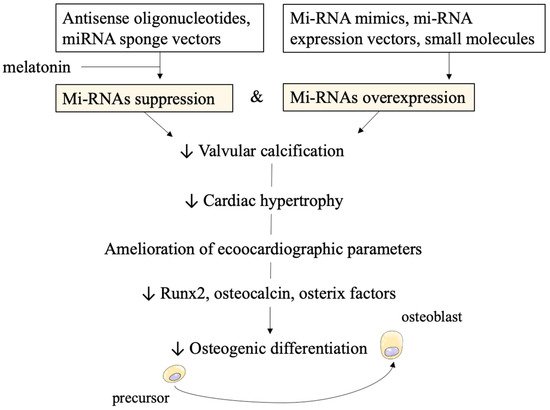
Another possible strategy recently proposed, involves melatonin administration. In vitro studies confirmed melatonin reduces the level of CircRIC3, a circular RNA with procalcific effects. It acts as a miR-204-5p sponge to stimulate and increase expression levels of the procalcification gene dipeptidyl peptidase-4 (DPP4). A preclinical in vivo study [
86] involving high cholesterol diet (HCD)-treated ApoE
−/− mice with aortic valve calcification demonstrated that the intragastric administration of melatonin for 24 weeks improved aortic valvular parameters. It reduced thickness and calcium deposition in the leaflets and ameliorated echocardiographic markers, namely transvalvular peak jet velocity and aortic valve area [
86]. At the molecular level, it decreased Runx2, osteocalcin, and osterix factors which are involved in osteogenic differentiation, as we discussed above (
Figure 2).
Figure 2. Possible therapeutic strategies and in vitro/in vivo effects of miRNAs targeting. Technological approaches of suppressing and overexpressing miRNAs are displayed at the top of the figure. The subsequent reduction in calcification clinical parameters and molecular pathways is achieved.
The authors also demonstrated melatonin caused in vitro suppression of calcification in human valvular interstitial cells (hIVICs) [
86].
3.2. Technical Concerns on Stability and Efficacy of miRNAs as Therapeutical Targets
The primary concerns in utilizing miRNAs as therapeutic targets, either to positively or to negatively affect them, arise from the need of achieving stability and resistance to degradation enzymes. We named expression vectors, antisense nucleotides (ASOs), small molecules and miR-mimics as novel approaches under current experimentation. Expression vectors are also defined as miRNAs sponges and constitute artificial binding sites for miRNAs to reduce their effect on miRNAs [
87,
88]. Oligonucleotides also bind miRNAs but are regarded as anti-miR for their sequence complementarity. With this strategy, they relieve miRNA targets from degradation or transcriptional blockage. Small molecules serve as translational regulators instead, but specific targets have not been revealed yet.
A possible strategy to improve stability is the modification with 2-O-methyl (2-OMe) [
89]. This can be then further stabilized with sulfur atoms in place of non-bridging oxygen atoms in the phosphate backbone. Serum nucleases, deputed to degradation of mi-RNAs, would find it difficult to cleave phosphorothioate bonds, given that they normally cleave phosphate bonds [
89,
90]. The adding of a 3′ cholesterol tail is another approach to ameliorate stability and efficacy.
To decrease nuclease degradation, modifications including 2-O-methoxyethyl (2-MOE), 2-fluoro (2-F), and locked nucleic acid (LNA) have also been tested [
84]. In particular, 2′-F-modifications yield resistant nucleotides only in combination with phosphorothioate modifications and proved to be the most effective one [
91].
3.3. Disadvantages in Pharmacokinetics and Proposed Mechanisms for Delivery Vehicles
In vitro studies conducted on oligonucleotides had limited pharmacological effects due to unfavorable kinetic characteristics, notably poor tissue distribution and fast excretion. Thus, appropriate delivery systems have been developed, functioning as carriers for in vivo molecular directing [
87].
A good delivery system should achieve the following features: evading the immune system response, avoiding nucleases degradation, directed to target cells, and releasing the content for incorporation into RNA processing machinery [
92,
93,
94,
95]. The main combination strategies include polymers, lipids, conjugation, antibodies, nanoparticles, and microbubbles [
87]. In particular, nanoparticles can deliver anti-miRNAs and small molecules with a greater degree of multi-functionality [
96]. The advantages of nanoparticles include large surface-to-volume ratios, hence, controlling their surface properties is crucial [
97]. Surface charges also appear fundamental: macrophage scavenging is increased when the charge increases in number (either positive or negative) [
96,
98,
99]. So, minimizing interactions to non-target sites via, as an instance, steric stabilization, would prevent nanoparticles from directing the molecules to undesired locations, and evading the immune system.
Other emerging technologies to improve kinetic parameters are nanoscale drug delivery systems using liposomes [
100]. Lower systemic toxicity has also been proved, especially in achieving high efficacies for anticancer therapies [
101,
102,
103]. Microbubbles, instead, have been used in combination with ultrasound to deliver anti-miRNAs after ischemia towards myocardiocytes of mice models. Molecular structure of microbubbles includes mixing of 1,2-distearoyl-sn-glycero-3-phosphocholine, 1,2-stearoyl-3-trimethylammonnium-propane and polyoxyethylene-40-stearate in H
2O, glycerol, and propylene glycol, in the presence of perfluorobutane gas [
87,
104,
105,
106,
107].
Local miRNA delivery results have been published and discussed in an Israeli study for metastatic breast cancer prevention by miR-96 and miR-182 treatment [
108]. In vivo, local targeting was achieved by coating breast tumor cells with adhesive hydrogel scaffold covered in nanoparticles carrying the miRNAs of interest [
108]. Nanoparticle stability achieved with hydrogel was also described for drug delivery in several other studies [
109,
110,
111].
This entry is adapted from the peer-reviewed paper 10.3390/ijms222212132