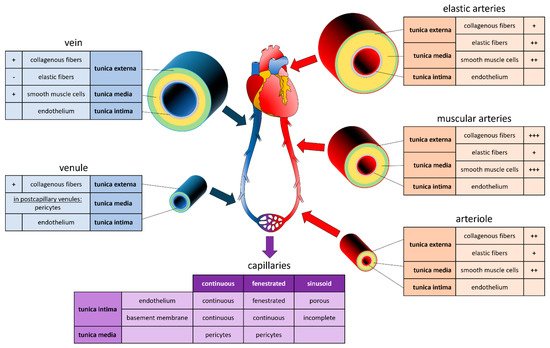
To fabricate blood vessels and vascularized tissue in vitro, it is critical to understand the biological, physiological, and functional aspects of the in vivo system. The peripheral vascular system (PVS) includes all the blood vessels exiting the heart. Blood vessels are classified in arteries, capillaries, or veins. Arteries transport the blood from the heart to the periphery. They branch into smaller and smaller arteries until they branch into arterioles, metarterioles, and capillaries, the smallest and most numerous blood vessels. The primary function of capillaries is to accommodate the exchange of nutrients, gases, and bi-products between the blood and the cells. They form the connection between arteries and veins, which transport the blood back towards the heart. Capillaries connect to venules merging into increasingly larger veins. It is worth noting that large segments of the PVS are identified as separated anatomical structures while microvasculature (arterioles, metarterioles, capillaries, and venules) are parts of the tissue they vascularized [
11,
12].
The vessel's structure varies according to the diameter of the vessels, which, in turn, is adapted to the hydrostatic pressure present in its interior (
Figure 1). Aside from capillaries, blood vessels are composed of three concentric layers, starting from the lumen they are: the tunica intima, the tunica media, and the adventitia. The tunica intima is composed of a monolayer of endothelial cells (EC) that repose on a basement membrane composed of type IV collagen and laminin. The outer layer of the tunica intima is composed of an elastin layer, also known as the
internal elastic lamina. The functions of the tunica intima are to contain cells and fluid within the vessel’s lumen and allow the blood to flow without problems. The tunica media is the middle layer of arteries and veins. It is primarily composed of smooth muscle cells (SMCs), collagen type I and type III, and variable amounts of elastic fibers, depending on the vessel thickness. It supports blood vessels and changes their diameter to regulate blood flow and blood pressure [
13]. The external elastic lamina separates the tunica media from the adventitia. The outer layer, the adventitia, is composed of a loose connective tissue of type I collagen and fibroblasts. Its primary function is to restrain the vessel from excessive extension and recoil. These three layers are present in the macro- and micro-vasculature of arteries and veins, except for the capillaries where SMCs are absent and replaced by pericytes. The thickness of the layers is inversely correlated with the degree of branching.
Figure 1. Composition of Vessels in the Cardiovascular System. Graphic and text inserts represent the composition of arteries (red), veins (blue) and capillaries (purple). For each section, the tunica externa (green), the tunica media (orange), and the tunica intima (grey) are indicated, as well as the proportion in collagenous fibers, elastic fibers, smooth muscle cells, and endothelium, ranging from lowest (-) to highest (+++) proportion.
Systemic arteries can be divided into two types, i.e., elastic and muscular, according to their tunica media’s relative composition in elastin and SMCs [
14]. Arteries with close proximity to the heart, like the aorta and the pulmonary arteries, are very elastic. Their tunica media contains more elastin than SMCs. This characteristic allows them to maintain a constant pressure gradient, despite the periodic changes in cardiac flow volume. Muscular arteries are medium-sized arteries (e.g., femoral arteries, brachial arteries, radial arteries) and contain more SMCs than elastin in the tunica media. This allows them to contract and expand to adjust their diameter according to the peripheral blood demand, thereby controlling the blood flow in the capillaries. Capillaries are thin-wall vessels with lumen diameters of about 5 to 10 µm. Their thin wall, composed of a single endothelium layer, allows the exchange of nutrients, metabolites, and gases primarily via diffusion. In addition, differences observed in the capillary structure allow different solute permeability in different organs. Accordingly, capillaries are classified as continuous, fenestrated, and sinusoid capillaries [
14]. Continuous capillaries have a continuous basement membrane, and their cells are connected by tight cell junctions. They are found in muscles, nervous systems, lungs, and skin [
15]. Fenestrated capillaries receive their name from window-like transcellular openings, that generate a mesh-like structure. They are found in exocrine glands, renal glomeruli, and intestinal mucosa [
15]. Finally, sinusoid capillaries have large interstitial gaps and an incomplete basement membrane. Sinusoid capillaries are found in the liver, spleen, and bone marrow [
15]. Several capillaries join to form a venule. Venules have a diameter of 8 to 100 µm, with few SMCs, little elastin, and a thin adventitia. Venules join to form veins. Like arteries, veins are composed of three layers. However, veins are thin-walled, with a reduced tunica media and less elastic. This particularity allows the venous system to accommodate a large volume of blood (~3/4 of the circulating blood) at relatively low pressure. Unlike arteries, veins contain valves that prevent backflow and allow the blood to reach the heart [
11].