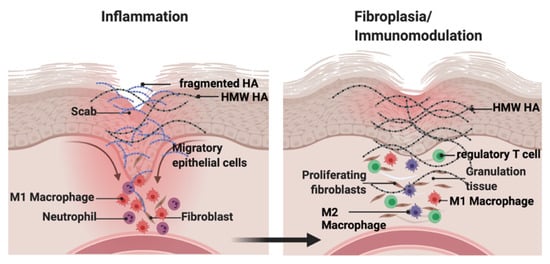
Hyaluronan (HA) is a major component of the ECM that consists of repeated disaccharide units of N-acetylglucosamine and glucuronic acid. HA is synthesized as a high molecular weight molecule but is degraded into heterogeneous fragments by hyaluronidases and reactive oxygen or nitrogen species. The biological activity of HA depends on its molecular weight. HA fragments stimulate tissue inflammation and fibrosis. In contrast, high molecular weight HA suppresses these processes and maintains tissue homeostasis. This functional duality is particularly important during wound repair where HA sequentially promotes then suppresses inflammation and fibrosis. In contrast, in tumors, HA effects are often co-opted to increase growth and invasion.
- hyaluronan
- RHAMM
- CD44
- wound repair
- tumor progression
- inflammation
- fibrosis
1. Background
2. The Hyaluronome
2.1. Hyaluronan
2.2. Hyaluronan Synthases
2.3. Hyaluronidases
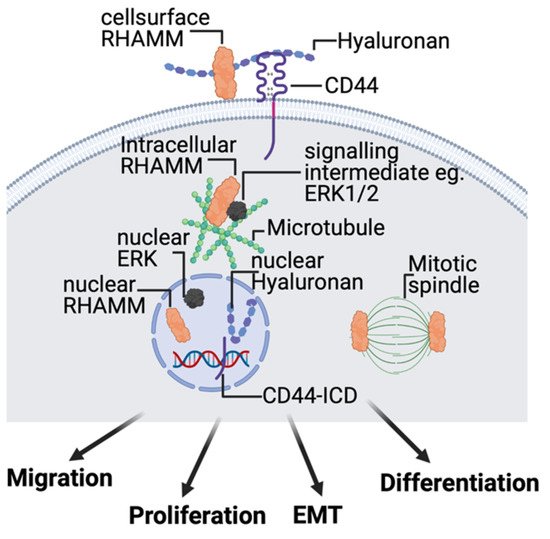
2.4. Hyaluronan Receptors, CD44, and RHAMM
This entry is adapted from the peer-reviewed paper 10.3390/biom11111551