Intrauterine growth restriction (IUGR) is associated with reduced placental amino acid transport (AAT). However, it remains to be established if changes in AAT contribute to restricted fetal growth. We hypothesized that reduced in vivo placental AAT precedes the development of IUGR in baboons with maternal nutrient restriction (MNR). Baboons were fed either a control (ad libitum) or MNR diet (70% of control diet) from gestational day (GD) 30. At GD 140, in vivo transplacental AA transport was measured by infusing nine (13)C- or (2)H-labeled essential amino acids (EAAs) as a bolus into the maternal circulation at cesarean section. A fetal vein-to-maternal artery mole percent excess ratio for each EAA was measured. Microvillous plasma membrane (MVM) system A and system L transport activity were determined. Fetal and placental weights were not significantly different between MNR and control. In vivo, the fetal vein-to-maternal artery mole percent excess ratio was significantly decreased for tryptophan in MNR. MVM system A and system L activity was markedly reduced in MNR. Reduction of in vivo placental amino acid transport precedes fetal growth restriction in the non-human primate, suggesting that reduced placental amino acid transfer may contribute to IUGR.
- maternal-fetal exchange
- amino acids
- neutral
- trophoblast
- fetal growth restriction
1. Introduction
2. Development and Findings
| Parameters | Gestation Day 120 | Gestation Day 140 | Gestation Day 165 |
|---|---|---|---|
| Reference | [34] | Current study | [11][36] |
| Fetal weight | ⬌ | ⬌ | ↓ |
| Placental weight | ⬌ | ⬌ | ↓ |
| MVM System A activity | ↓ | ↓ | ↓ |
| MVM System A amino acid transporter isoforms (SNAT1,2 and 4) expression | ⬌ | ↓SNAT2 | ↓SNAT2; ⬌ SNAT1, SNAT4 |
| MVM System L activity | ⬌ | ↓ | ↓ |
| MVM System L amino acid transporter isoforms expression (LAT1 and 2) | ⬌ | ↓LAT1 | ↓LAT1 and LAT2 |
| MVM Taurine transporter expression | ⬌ | ⬌ | ↓ |
| BM System L activity | ⬌ | ↓ | ↓ |
| BM System L amino acid transporter LAT1 isoform expression | ⬌ | Not studied | Not studied |
| Maternal plasma concentration of amino acids | ⬌ | Not studied | ↓Aspartic acid, glutamic acid, tyrosine, tryptophan, phenylalanine, leucine, and ornithine; ↑Glycine |
| Fetal plasma concentration of amino acids | ↓ Leucine and isoleucine. ↑Citrulline |
Not studied | ↓Taurine, tyrosine, phenylalanine, leucine, and ornithine |
| In vivo transplacental amino acid transport | Not studied | ↓Tryptophan | ↓Leucine, isoleucine, methionine, phenylalanine, threonine, and tryptophan |
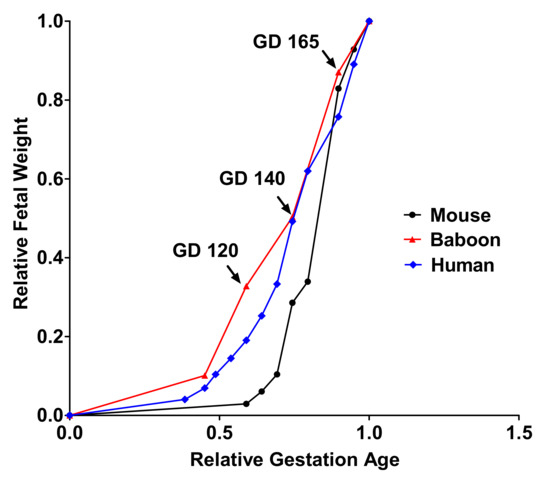
3. Conclusions
This entry is adapted from the peer-reviewed paper 10.3390/nu13082892
References
- Malhotra, A.; Allison, B.J.; Castillo-Melendez, M.; Jenkin, G.; Polglase, G.R.; Miller, S.L. Neonatal Morbidities of Fetal Growth Restriction: Pathophysiology and Impact. Front Endocrinol. 2019, 10, 55.
- Sharma, D.; Shastri, S.; Farahbakhsh, N.; Sharma, P. Intrauterine growth restriction—Part 1. J. Matern Fetal Neonatal Med. 2016, 29, 3977–3987.
- Brodsky, D.; Christou, H. Current concepts in intrauterine growth restriction. J. Intensive Care Med. 2004, 19, 307–319.
- Gluckman, P.D.; Hanson, M.A.; Cooper, C.; Thornburg, K.L. Effect of in utero and early-life conditions on adult health and disease. N. Engl. J. Med. 2008, 359, 61–73.
- Hales, C.N.; Barker, D.J.; Clark, P.M.; Cox, L.J.; Fall, C.; Osmond, C.; Winter, P.D. Fetal and infant growth and impaired glucose tolerance at age 64. BMJ 1991, 303, 1019–1022.
- Barker, D.J.; Gluckman, P.D.; Godfrey, K.M.; Harding, J.E.; Owens, J.A.; Robinson, J.S. Fetal nutrition and cardiovascular disease in adult life. Lancet 1993, 341, 938–941.
- Sibley, C.P.; Turner, M.A.; Cetin, I.; Ayuk, P.; Boyd, C.A.; D’Souza, S.W.; Glazier, J.D.; Greenwood, S.L.; Jansson, T.; Powell, T. Placental phenotypes of intrauterine growth. Pediatr. Res. 2005, 58, 827–832.
- Nord, M.; Andrews, C.J.; Andrews, M.S.; Carlson, S. Household Food Security in the United States 2009; United States Department of Agriculture, Economic Research Services: Washington, DC, USA, 2020.
- Jansson, N.; Pettersson, J.; Haafiz, A.; Ericsson, A.; Palmberg, I.; Tranberg, M.; Ganapathy, V.; Powell, T.L.; Jansson, T. Down-regulation of placental transport of amino acids precedes the development of intrauterine growth restriction in rats fed a low protein diet. J. Physiol. 2006, 576, 935–946.
- Roos, S.; Jansson, N.; Palmberg, I.; Saljo, K.; Powell, T.L.; Jansson, T. Mammalian target of rapamycin in the human placenta regulates leucine transport and is down-regulated in restricted fetal growth. J. Physiol. 2007, 582, 449–459.
- Kavitha, J.V.; Rosario, F.J.; Nijland, M.J.; McDonald, T.J.; Wu, G.; Kanai, Y.; Powell, T.L.; Nathanielsz, P.W.; Jansson, T. Down-regulation of placental mTOR, insulin/IGF-I signaling, and nutrient transporters in response to maternal nutrient restriction in the baboon. FASEB J. 2014, 28, 1294–1305.
- Rosario, F.J.; Jansson, N.; Kanai, Y.; Prasad, P.D.; Powell, T.L.; Jansson, T. Maternal protein restriction in the rat inhibits placental insulin, mTOR, and STAT3 signaling and down-regulates placental amino acid transporters. Endocrinology 2011, 152, 1119–1129.
- Verrey, F. System L: Heteromeric exchangers of large, neutral amino acids involved in directional transport. Pflug. Arch. 2003, 445, 529–533.
- del Amo, E.M.; Urtti, A.; Yliperttula, M. Pharmacokinetic role of L-type amino acid transporters LAT1 and LAT2. Eur. J. Pharm. Sci. 2008, 35, 161–174.
- Gaccioli, F.; Aye, I.L.; Roos, S.; Lager, S.; Ramirez, V.I.; Kanai, Y.; Powell, T.L.; Jansson, T. Expression and functional characterisation of System L amino acid transporters in the human term placenta. Reprod. Biol. Endocrinol. 2015, 13, 57.
- Christensen, H.N.; Oxender, D.L.; Liang, M.; Vatz, K.A. The use of N-methylation to direct route of mediated transport of amino acids. J. Biol. Chem. 1965, 240, 3609–3616.
- Matoba, S.; Nakamuta, S.; Miura, K.; Hirose, M.; Shiura, H.; Kohda, T.; Nakamuta, N.; Ogura, A. Paternal knockout of Slc38a4/SNAT4 causes placental hypoplasia associated with intrauterine growth restriction in mice. Proc. Natl. Acad. Sci. USA 2019, 116, 21047–21053.
- Chen, Y.Y.; Rosario, F.J.; Abu Shehab, M.; Powell, T.L.; Gupta, M.B.; Jansson, T. Increased ubiquitination and reduced plasma membrane trafficking of placental amino acid transporter SNAT-2 in human IUGR. Clin. Sci. 2015, 129, 1131–1141.
- Jansson, T.; Ylven, K.; Wennergren, M.; Powell, T.L. Glucose transport and system A activity in syncytiotrophoblast microvillous and basal plasma membranes in intrauterine growth restriction. Placenta 2002, 23, 392–399.
- Shibata, E.; Hubel, C.A.; Powers, R.W.; von Versen-Hoeynck, F.; Gammill, H.; Rajakumar, A.; Roberts, J.M. Placental system A amino acid transport is reduced in pregnancies with small for gestational age (SGA) infants but not in preeclampsia with SGA infants. Placenta 2008, 29, 879–882.
- Mahendran, D.; Donnai, P.; Glazier, J.D.; D’Souza, S.W.; Boyd, R.D.; Sibley, C.P. Amino acid (system A) transporter activity in microvillous membrane vesicles from the placentas of appropriate and small for gestational age babies. Pediatr. Res. 1993, 34, 661–665.
- Glazier, J.D.; Cetin, I.; Perugino, G.; Ronzoni, S.; Grey, A.M.; Mahendran, D.; Marconi, A.M.; Pardi, G.; Sibley, C.P. Association between the activity of the system A amino acid transporter in the microvillous plasma membrane of the human placenta and severity of fetal compromise in intrauterine growth restriction. Pediatr. Res. 1997, 42, 514–519.
- Mando, C.; Tabano, S.; Pileri, P.; Colapietro, P.; Marino, M.A.; Avagliano, L.; Doi, P.; Bulfamante, G.; Miozzo, M.; Cetin, I. SNAT2 expression and regulation in human growth-restricted placentas. Pediatr. Res. 2013, 74, 104–110.
- Rodriguez, J.S.; Zurcher, N.R.; Keenan, K.E.; Bartlett, T.Q.; Nathanielsz, P.W.; Nijland, M.J. Prenatal betamethasone exposure has sex specific effects in reversal learning and attention in juvenile baboons. Am. J. Obs. Gynecol. 2011, 204, 545-e1.
- Zurcher, N.R.; Rodriguez, J.S.; Jenkins, S.L.; Keenan, K.; Bartlett, T.Q.; McDonald, T.J.; Nathanielsz, P.W.; Nijland, M.J. Performance of juvenile baboons on neuropsychological tests assessing associative learning, motivation and attention. J. Neurosci. Methods 2010, 188, 219–225.
- Nijland, M.J.; Schlabritz-Loutsevitch, N.E.; Hubbard, G.B.; Nathanielsz, P.W.; Cox, L.A. Non-human primate fetal kidney transcriptome analysis indicates mammalian target of rapamycin (mTOR) is a central nutrient-responsive pathway. J. Physiol. 2007, 579, 643–656.
- Kamat, A.; Nijland, M.J.; McDonald, T.J.; Cox, L.A.; Nathanielsz, P.W.; Li, C. Moderate global reduction in maternal nutrition has differential stage of gestation specific effects on 1- and 2-adrenergic receptors in the fetal baboon liver. Reprod. Sci. 2011, 18, 398–405.
- Schlabritz-Loutsevitch, N.E.; Dudley, C.J.; Gomez, J.J.; Nevill, C.H.; Smith, B.K.; Jenkins, S.L.; McDonald, T.J.; Bartlett, T.Q.; Nathanielsz, P.W.; Nijland, M.J. Metabolic adjustments to moderate maternal nutrient restriction. Br. J. Nutr. 2007, 98, 276–284.
- Cox, L.A.; Schlabritz-Loutsevitch, N.; Hubbard, G.B.; Nijland, M.J.; McDonald, T.J.; Nathanielsz, P.W. Gene expression profile differences in left and right liver lobes from mid-gestation fetal baboons: A cautionary tale. J. Physiol. 2006, 572, 59–66.
- Nijland, M.J.; Mitsuya, K.; Li, C.; Ford, S.; McDonald, T.J.; Nathanielsz, P.W.; Cox, L.A. Epigenetic modification of fetal baboon hepatic phosphoenolpyruvate carboxykinase following exposure to moderately reduced nutrient availability. J. Physiol. 2010, 588, 1349–1359.
- Cox, L.A.; Nijland, M.J.; Gilbert, J.S.; Schlabritz-Loutsevitch, N.E.; Hubbard, G.B.; McDonald, T.J.; Shade, R.E.; Nathanielsz, P.W. Effect of 30 per cent maternal nutrient restriction from 0.16 to 0.5 gestation on fetal baboon kidney gene expression. J. Physiol. 2006, 572, 67–85.
- Rodriguez, J.S.; Zurcher, N.R.; Bartlett, T.Q.; Nathanielsz, P.W.; Nijland, M.J. CANTAB delayed matching to sample task performance in juvenile baboons. J. Neurosci. Methods 2011, 196, 258–263.
- Antonow-Schlorke, I.; Schwab, M.; Cox, L.A.; Li, C.; Stuchlik, K.; Witte, O.W.; Nathanielsz, P.W.; McDonald, T.J. Vulnerability of the fetal primate brain to moderate reduction in maternal global nutrient availability. Proc. Natl. Acad. Sci. USA 2011, 108, 3011–3016.
- Pantham, P.; Rosario, F.J.; Weintraub, S.T.; Nathanielsz, P.W.; Powell, T.L.; Li, C.; Jansson, T. Down-Regulation of Placental Transport of Amino Acids Precedes the Development of Intrauterine Growth Restriction in Maternal Nutrient Restricted Baboons. Biol. Reprod. 2016, 95, 98.
- Van Calsteren, K.; de Catte, L.; Devlieger, R.; Chai, D.C.; Amant, F. Sonographic biometrical normograms and estimation of fetal weight in the baboon (Papio anubis). J. Med. Primatol. 2009, 38, 321–327.
- Pantham, P.; Rosario, F.J.; Nijland, M.; Cheung, A.; Nathanielsz, P.W.; Powell, T.L.; Galan, H.L.; Li, C.; Jansson, T. Reduced placental amino acid transport in response to maternal nutrient restriction in the baboon. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2015, 309, R740–R746.
- Ma, Y.; Zhu, M.J.; Uthlaut, A.B.; Nijland, M.J.; Nathanielsz, P.W.; Hess, B.W.; Ford, S.P. Upregulation of growth signaling and nutrient transporters in cotyledons of early to mid-gestational nutrient restricted ewes. Placenta 2011, 32, 255–263.
- Jansson, T.; Scholtbach, V.; Powell, T.L. Placental transport of leucine and lysine is reduced in intrauterine growth restriction. Pediatr. Res. 1998, 44, 532–537.
- Ganapathy, M.E.; Leibach, F.H.; Mahesh, V.B.; Howard, J.C.; Devoe, L.D.; Ganapathy, V. Characterization of tryptophan transport in human placental brush-border membrane vesicles. Biochem. J. 1986, 238, 201–208.
- Kudo, Y.; Boyd, C.A. Characterisation of L-tryptophan transporters in human placenta: A comparison of brush border and basal membrane vesicles. J. Physiol. 2001, 531, 405–416.
- Sano, M.; Ferchaud-Roucher, V.; Kaeffer, B.; Poupeau, G.; Castellano, B.; Darmaun, D. Maternal and fetal tryptophan metabolism in gestating rats: Effects of intrauterine growth restriction. Amino Acids 2016, 48, 281–290.
- Xu, K.; Liu, G.; Fu, C. The Tryptophan Pathway Targeting Antioxidant Capacity in the Placenta. Oxid Med. Cell Longev. 2018, 2018, 1054797.
- Huber, H.F.; Ford, S.M.; Bartlett, T.Q.; Nathanielsz, P.W. Increased aggressive and affiliative display behavior in intrauterine growth restricted baboons. J. Med. Primatol. 2015, 44, 143–157.
- Teodoro, G.F.; Vianna, D.; Torres-Leal, F.L.; Pantaleao, L.C.; Matos-Neto, E.M.; Donato, J., Jr.; Tirapegui, J. Leucine is essential for attenuating fetal growth restriction caused by a protein-restricted diet in rats. J. Nutr. 2012, 142, 924–930.
- Gaccioli, F.; Lager, S.; Powell, T.L.; Jansson, T. Placental transport in response to altered maternal nutrition. J. Dev. Orig. Health Dis. 2013, 4, 101–115.
- Rosso, P. Maternal-fetal exchange during protein malnutrition in the rat. Placental transfer of glucose and a nonmetabolizable glucose analog. J. Nutr. 1977, 107, 20006–20010.
- Rosso, P. Maternal-fetal exchange during protein malnutrition in the rat. Placental transfer of alpha-amino isobutyric acid. J. Nutr. 1977, 107, 2002–2005.
- Rosso, P. Maternal malnutrition and placental transfer of alpha-aminoisobutyric acid in the rat. Science 1975, 187, 648–650.
- Varma, D.R.; Ramakrishnan, R. Effects of protein-calorie malnutrition on transplacental kinetics of aminoisobutyric acid in rats. Placenta 1991, 12, 277–284.
- Sferruzzi-Perri, A.N.; Vaughan, O.R.; Coan, P.M.; Suciu, M.C.; Darbyshire, R.; Constancia, M.; Burton, G.J.; Fowden, A.L. Placental-specific Igf2 deficiency alters developmental adaptations to undernutrition in mice. Endocrinology 2011, 152, 3202–3212.
- Coan, P.M.; Vaughan, O.R.; Sekita, Y.; Finn, S.L.; Burton, G.J.; Constancia, M.; Fowden, A.L. Adaptations in placental phenotype support fetal growth during undernutrition of pregnant mice. J. Physiol. 2010, 588, 527–538.
- Coan, P.M.; Vaughan, O.R.; McCarthy, J.; Mactier, C.; Burton, G.J.; Constancia, M.; Fowden, A.L. Dietary composition programmes placental phenotype in mice. J. Physiol. 2011, 589, 3659–3670.