Torulaspora delbrueckii has attracted interest in recent years, especially due to its biotechnological potential, arising from its flavor- and aroma-enhancing properties when used in wine, beer or bread dough fermentation, as well as from its remarkable resistance to osmotic and freezing stresses.
- Torulaspora delbrueckii
- non-Saccharomyces
- wine
- bread
- biotechnology
- genomics
1. Introduction
2. Occurrence and General Characteristics
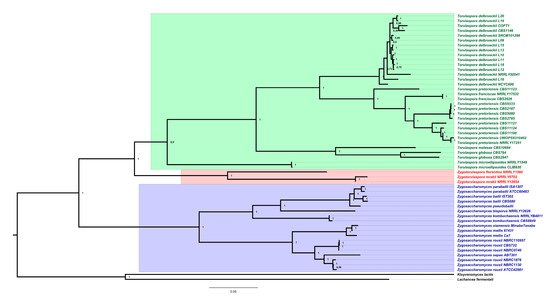
3. Genomics and Taxonomy
4. Metabolism
|
Product |
Torulaspora delbrueckii |
Saccharomyces cerevisiae |
Notes |
References |
|---|---|---|---|---|
|
Acetic acid |
0.27–0.56 g/L |
1.0–1.17 g/L |
Key signature in volatile acidity of wines |
|
|
Malic acid |
Consumption between 10.5–25% |
Whether degradation or production is desirable depends on the must characteristics. |
||
|
Citric acid |
2.18–2.36 g/L |
2.23 g/L |
Citrus-like taste |
[41] |
|
Succinic acid |
0.84–1.11 g/L |
Maximum of 0.65 g/L |
Minor acid in the overall wine acidity, although the combination with one molecule of ethanol creates the ester mono-ethyl succinate, responsible for a mild, fruity aroma |
[43] |
|
- |
Maximum of 1.13 g/L |
[44] |
||
|
Mannoproteins |
T. delbrueckii produces 25% more than S. cerevisiae |
Released during fermentation or ageing processes |
[23] |
|
|
Polysaccharides |
T. delbrueckii releases 50% more than S. cerevisiae |
Increases the quality of mouthfeel properties |
[45] |
|
|
Glycerol |
1–10.5 g/L |
Maximum of 9.1 g/L |
Smoothness and viscosity features |
|
|
Ethanol |
40.6–72.68 g/L |
103–121 g/L |
||
5. Biotechnological Importance of T. delbrueckii
5.1. Bread Industry
5.2. Production of Fermented Beverages
|
Beverages Applications |
Used Substrate |
Advantages |
Disadvantages |
References |
|---|---|---|---|---|
|
Beer |
Wort |
High tolerance to hop compounds; good flavor-forming properties |
Low sugar utilization |
|
|
Mezcal |
Agave juice † |
Rich in volatile compounds; acceptable in sensory tests |
Low performance |
|
|
Tequila |
Agave juice * |
Positive influence on the final sensory profile |
– |
[74] |
|
Cider |
Apple juice † |
Great production of ethyl decanoate and ethyl hexanoate |
Low performance; negligible amounts of acetate esters |
|
|
Mead |
Honey sugar |
Good fermentation ability; Good sensory features |
Grassy flavor |
[7] |
|
Soy alcoholic beverage |
Soy whey |
Enrich aroma profiles: high levels of ethyl decanoate and ethyl hexanoate; metabolize hexanal; |
– |
[75] |
* Specifically from Agave tequilana; † sterile.
5.3. Other Food Applications
|
Food Applications |
Used Substrate |
Advantages |
Disadvantages |
References |
|---|---|---|---|---|
|
Chocolate |
Cocoa beans |
Good flavor quality of cocoa and, therefore, the chocolate |
Expedite in mixed fermentations with S. cerevisiae |
[77] |
|
Cheese |
Cheese |
Varied aromatic properties |
Unable to inhibit pathogenic bacteria; low β-glucosidase activity |
|
|
Honey |
Honey sugar |
Rapidly ferment sugar |
Large-scale productions only in combination with S. cerevisiae |
[7] |
|
Olive oil |
Black olives |
Easy hydrolyzation of olive oil |
Growth inhibition at concentrations higher than 0.5% (w/v) of oleuropein |
[81] |
|
Coffee |
Coffee cherries |
Improve coffee’s sensorial quality |
Pronounced astringency depending on the coffee variety |
|
|
Bio-protection |
– |
Reduction in the use of chemical preservatives to control food spoilage |
– |
This entry is adapted from the peer-reviewed paper 10.3390/jof7090712