Algal species still show unrevealed and unexplored potentiality for the identification of new compounds. Photosynthetic organisms represent a valuable resource to exploit and sustain the urgent need of sustainable and green technologies. Particularly, unconventional organisms from extreme environments could hide properties to be employed in a wide range of biotechnology applications, due to their peculiar alleles, proteins, and molecules. In this review we report a detailed dissection about the latest and advanced applications of protein derived from algae. Furthermore, the innovative use of modified algae as bio-reactors to generate proteins or bioactive compounds was discussed. The latest progress about pharmaceutical applications, including the possibility to obtain drugs to counteract virus (as SARS-CoV-2) were also examined.
- microalgae
- diatoms
- biodiesel
- synthetic biology
- extreme environments
- recombinant protein
- vaccines
1. Introduction
In recent years, public opinion, research community, and commercial stakeholders paid great attention to green and sustainable biotechnologies; these are not anymore considered a luxury, but a necessity in all countries. Actually, the current world population is estimated in 7.8 billion people, and an increase is expected of about 2 billions in the next 30 years (UN; www.un.org ; accessed on 1 July 2021); therefore, it is essential to develop sustainable biotechnologies and strategies to face the increased food demand with a reduced impact on natural environment. Plants and algae possess still untouched possibilities; thus, these organisms are emerging as formidable sustainable tools with a number of advantages over the conventional biotechnologies [1,2,3,4].
Since 1980s, plants emerged as one of the most promising production platforms for recombinant proteins and for the valuable production of bio-compounds [5,6]. The agricultural scale cultivation represents a cost-effective method to produce recombinant proteins as well as an innovative approach to reduce risks of product contamination [2]. Encouraging results in the production of recombinant proteins and molecules for pharmaceutical applications were preliminarily obtained using higher plants [7]. On the other hand, the intensive exploitation of cropland represents a severe constraint influencing the environment utilization, CO 2 emission, chemical pollution, and water quality and availability [8].
At the same time, microalgae overcome a number of limitations of plants, emerging as effective biotechnological platforms and giving added value by the exploitation of bioactive compounds obtained from these biomasses [9,10]. Microalgae are characterized for being unicellular and their versatile metabolism, representing efficient and economic platforms to gather organics compounds such as proteins, lipids, pigments, sterols, and carbohydrates for a number of commercial applications such as nutraceutical, pharmaceutical and for biofuels [11,12]. This group includes photosynthetic prokaryotes (e.g., cyanobacteria) or eukaryotes, which are able to live in diversified environments [9]. As consequence, a number of microalgae species are currently cultivated for commercial and industrial aims, and more than 75% of this production is related to healthier supplements for human consumption [10].
In this review, we analyze some recent examples about the utilization of photosynthetic organisms from marine environments, as bio-factories. Furthermore, we reported a survey of the latest and advanced applications of proteins and bioactive compounds from microalgae.
2. Microalgae Engineering to Obtain Platforms for Biocompounds Productions
Thus, a number of strategies have been used to hyperaccumulate fatty acids and/or triacylglycerol (TAG) content ( Table 1 ). Phaeodactylum tricornutum and Thalassiosira pseudonana engineered strains were obtained modifying TAG biosynthetic pathways. In this context, critical enzymes are glycerol-3-phosphate acyltransferase (GPAT), lysophosphatidic acid acyl- transferase (LPAAT), and acyl-CoA:diacylglycerol acyltransferase (DGAT). These enzymes regulate fatty acid and TAG biosynthetic pathways in plants and microalgae [22,23,24]. The overexpression of GPAT is effective to obtain suitable strains for biofuel production ( Table 1 ). Both isoforms GPAT1 and GPAT2 were overexpressed in P. tricornutum resulting in a 2.9- and 2.3-fold change increased lipid content, respectively [19,20]. Interestingly, the overexpression of GPAT2 regulated the natural expression of LPAT and DGAT, causing an enhanced effect on total lipid content [20]. Interestingly, this strain was also reported as better tolerant to abiotic stress, namely salinity and chilling [20]. A similar approach was recently used in Neochloris oleoabundans where the plastidial Neo LPAAT1 and endoplasmic reticulum-located Neo DGAT2 were co-overexpressed. This strategy increased the TAG content of about 2-fold compared to wild type; furthermore, engineered strain reporting no effects on algal biomass [25]. Significant results were obtained using heterologous expression, mutagenesis in situ and knockdown strategies of genes related to the lipid biosynthetic pathway. Trentecoste et al. [26] and Barka et al. [27], developed specific lipase knockout strains of T. pseudonana and P. tricornutum, respectively. The suppression of TAG lipase in these two diatoms significantly increased the lipid content. Interestingly both did not show effects on growth [12,26,27]. TALEN-based genome editing technique was used in P. tricornutum strain [28] reporting modification in Pt TES1, a hotdog-fold thioesterase involved in acyl-CoA hydrolysis. This modification lead to a 1.7-fold change increase of TAG content [28]. Successful editing results were obtained by using CRISP/CAS9 approach on P. tricornutum and T. pseudonana . These preliminary results opened new perspectives to obtain improved microalgae strains using this technique [29,30,31].
An increase in lipids content has been obtained by modifying different microalgal pathways. LEAFY COTYLEDON1 from Arabidopsis thalian (At LEC1) represents a key transcription factor involved in plants lipid metabolism [32]. Liu et al. [33] demonstrated that the endogenous expression of At LEC1 improves fatty acids and total lipids content in Chlorella ellipsoidea both under mixotrophic and autotrophic culture conditions. These differences between wild type and the engineered algal strain were related to an enhanced regulation of key enzymes namely ACCase, GPDH, PDAT1, and DGAT1 [33]. Another interesting example of the transfer of genes from A. thaliana in microalgae showed the heterologous overexpression of 1-deoxy-D-xylulose 5-phosphate synthase from A. thaliana in Nannochloropsis oceanica . This strain reported an improved CO 2 fixation, thus increasing biomass, lipids, and carbohydrates productions upon different nutritional and stressed conditions [34]. Particularly, Nannochloropsis represented a further model used as platform for biofuel production from microalgae [34,35,36,37]. In the last year a number of papers have been published about the genetic manipulation of Nannochloropsis , particularly focused on the increase on lipid and fatty acids content. Different approaches, namely overexpression [36], endogenous expression [37], and insertional mutagenesis [38] were successfully used in Nannochloropsis . For example, Jeon et al. [36] overexpressed the NADP-dependent malic enzyme ( Ns ME1) in Nannochloropsis salina. This enzyme is an important NADPH supplier, playing the central role in the C4-like cycle of microalgae. The overexpressed strain showed an increased fatty acid and lipids contents compared with wild type, up to 53% and 38%, respectively [36].
Analogous to phycoerythrin, astaxanthin is a desirable product for human consumption. This is mainly produced by the green microalga Haematococcus pluvialis in response to adverse conditions namely excess of light, salinity, and nutrient starvation [51]. A number of mutagenesis approaches were used on H. pluvialis using physical and chemical mutagens such as UV radiation, ethyl methanesulphonate (EMS), diethyl sulphate (DES) and other. In some case desirable improved strains were obtained showing high growth rates at the vegetative stage and high astaxanthin accumulation rates at the encystment stage [51]. In recent years, genetic engineering approaches were also obtained in H. pluvialis . For example, Waissman-Levy et al. [52], manipulated the nuclear genome of H. pluvialis by insertion of the hexose uptake protein (HUP1) gene from the green microalga Parachlorella kesslerii . The engineered strain was able to grow upon heterotrophy conditions in glucose-supplemented media [52].

3. Microalgae Engineering for the Production of Pharmacological Proteins
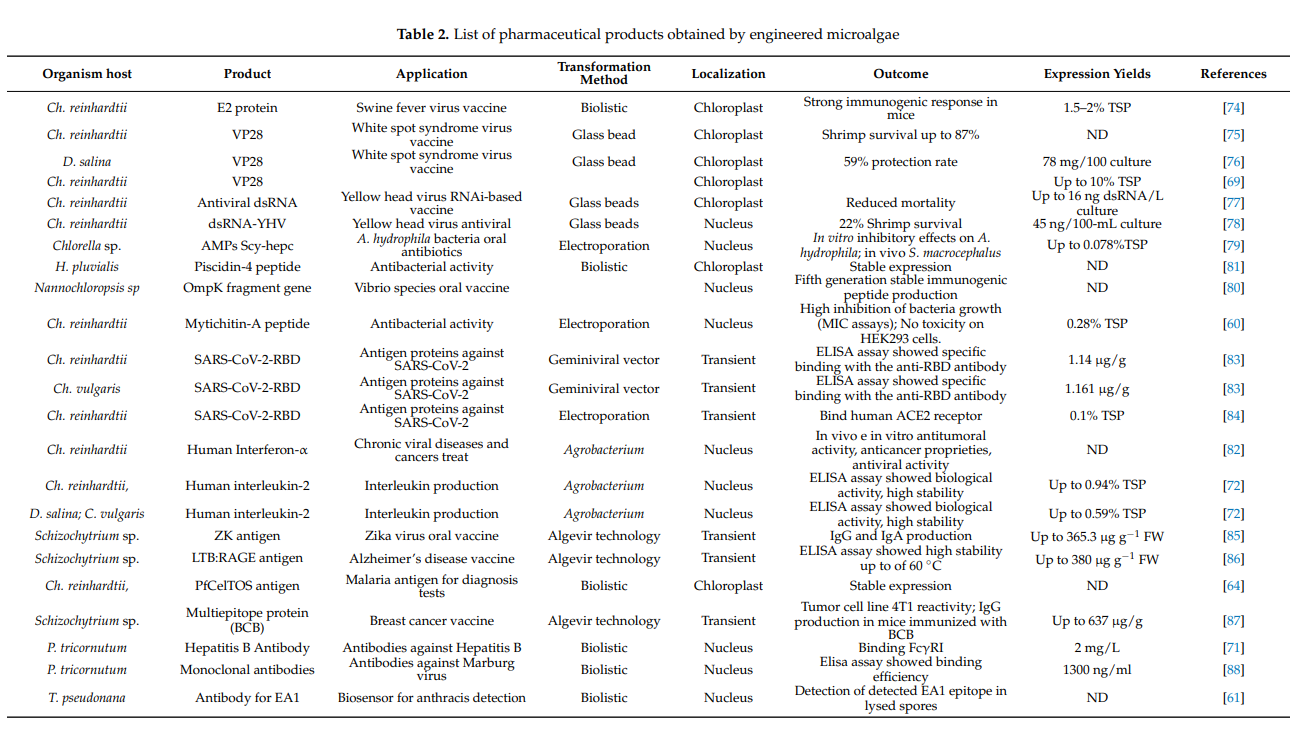
Microalgae represent an important biotechnology resource for the production of recombinant proteins for pharmacological applications. Successful results were reported by the genetic manipulation of microalgae for the production of antigens for vaccines, antibodies, immunotoxins, hormones, and antimicrobial agents [59,60,61,62,63,64,65,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87]. Strategies and techniques have been developed for the exploitation of microalgae as expression systems for a number of advanced genetic toolkits [62,63,64,65].
Recombinant protein production was attained by engineering both nuclear and chloroplast microalgal genomes [66]. Chloroplast manipulation produces a high level of transgene expression [66,67,68,69,70,71], but the recombinant proteins obtained could be not always subjected to critical post-translation modifications (PTMs), thus affecting their activity and functionality [63]. On the other hand, nuclear transformation produced a lower protein accumulation, but retained PTMs, such as N-glycosylation [59,67]. Furthermore, nuclear manipulation provided tools for the targeting of proteins into specific subcellular compartments or secreting them into culture media [72] while regulatory elements (promoters and terminators) and selectable markers were tested to identify improved strategies for higher levels of recombinant protein [15,73].
Recently, the utilization of different engineering strategies, many biopharmaceutical compounds have been effectively produced in microalgae, and Chlamydomonas reinhardtii has been widely used for this type of applications.
List of pharmaceutical products obtained by engineered microalgae (Table 2)
4. Extremophilic Microalgae as Bioreactors
A renewed attention has been focused on organisms from extreme environments [93,94]. This is particularly true for microalgae, which show a wide range survival capabilities under extreme and stress conditions, namely hypersalinity, high and low temperatures or toxic heavy metals levels [94,95,96,97]. The use of extremophilic microalgae showed notable benefits for various applications [93,98]. Commercial cultivations of microalgae are usually obtained by growth in photobioreactors and open ponds [54,98]. On the other hand, industrial biotechnology requires elevated temperatures, difficult sterilization procedures, and expensive downstream processing which benefit from the use of thermophilic microalgae [99,100]. Extreme operating conditions tolerate by specific microalgae strains were proposed to improve these industrial processes, thus overcoming contaminations, loss of biomass, meteorological constraints, and others [94,101,102]. Example of biotechnological applications are the cultivation of Chlorella to mitigate the effects of industrial pollution, by absorbing CO 2; the use of Arthrospira platensis for bioremediation of contaminated effluents; the utilization of a number of microalgal species for biofuel productions [100].
Thermophilic microalgae cultivated in large-scale open ponds are able to produce bulk amount of lipids, which can be utilised for biodiesel production technology [103]. Particularly, in recent years increasing attention were posed on the regulation of fatty acid dehydrogenase (FAD) in freeze resistant microalgae [59,104,105]. This studies contributed to both elucidation of the regulation of algal membrane assembly, and fatty acid metabolism. Promising results were obtained using the Δ12FAD from Chlamydomonas sp. ICE-L [59] and the Δ5FAD from Lobosphaera incise [105]. Both enzymes showed cold-resistant mechanisms to adapt their biochemical functions. Δ12FAD from Chlamydomonas sp. ICE-L contributes to membrane fluidity for adaptation to Antarctic extreme environment; Δ5FAD contributes to arachidonic acid synthesis, thus avoiding the risk of photodamage upon chilling. A number of snow algae were tested for their ability to produce lipid species in low temperatures [106]. These analysis reported an enriched production of unsaturated fatty acyl chains, especially C18:1n-9 and C18:3n-3, thus indicating these species as good candidates to improve the yields of microalgal biomass and oil products at low temperatures [106].
Thermophillic microalgae are studied for the applications of phycobiliproteins [107], a group of water-soluble proteins linked to chromophores, involved in light-harvesting processes. Phycobiliproteins find use in a wide range of commercial purposes (e.g., colorant for food and textile industries) and the exploitation of these proteins would give additional value [97]. A number of industrial processes require temperatures exceeding the stability of proteins from mesophiles [107]. Phycocianins from the cyanobacterium A. platensis are actually produced as dye in food industries but high temperatures (62 °C) limited their use [108]. Different authors proposed the use of phycocianins from the thermoacidophilic red alga Cyanidioschyzon merolae to overcome this problem [97,109]. The temperatures and pH tolerability of phycocianins from Cyanidioschyzon merolae were recently characterized by Yoshida et al. [96], confirming their thermotolerant properties. Similar evaluations were reported for phycobiliproteins in textile industry where the use of mesophiles proteins at high temperatures resulted in a decrease of color intensity [97].
A number of extromophilic microalgae were recently characterized for the ability in the production of bio-compounds with high economic interest [98]. For example, the snow green algae namely Chlamydomonas nivalis , Raphidonema sp., and Chloromonas sp. showed ability in the production of astaxanthin, α-Tocopherol, xanthophylles, and glycerol [110,111,112]. On the other hand, the heat tolerant green alga Desmodesmus and the acidophilic red alga Galdieria sulphuraria were tested for the production of lutein and blue pigment phycocianin, respectively [113,114]. These results demonstrate the unexplored potential represented by microalgae from extreme environments, thus encouraging the characterization of new species.
This entry is adapted from the peer-reviewed paper 10.3390/plants10081686