Noncommunicable diseases, such as Alzheimer’s disease, breast and prostate cancer, are becoming increasingly prevalent in Western countries. To better elucidate the onset and evolution of these pathologies and ultimately design new preventive and therapeutic strategies, research activities focused on these biomedical areas have been supported by the European Union in the last two decades. While research has globally contributed increasing our understanding of the pathological mechanisms underlying these diseases, the failure rate in drug development still remains very high. Nowadays, it is important to monitor contribution to innovation and impact of funded research by means of defined indicators.
- biomedical research
- Alzheimer’s disease
- breast cancer
- prostate cancer
- funding
- indicators
- translational failure
- animal models
- cross-disciplinarity.
1. Definition
Dementia and cancer are becoming increasingly prevalent in Western countries. In the last two decades, research focused on Alzheimer’s disease (AD) and cancer, in particular, breast cancer (BC) and prostate cancer (PC), has been substantially funded both in Europe and worldwide. While scientific research outcomes have contributed to increase our understanding of the disease etiopathology, still the prevalence of these chronic degenerative conditions remains very high across the globe. By definition, no model is perfect. In particular, animal models of AD, BC, and PC have been and still are traditionally used in basic/fundamental, translational, and preclinical research to study human disease mechanisms, identify new therapeutic targets, and develop new drugs. However, animals do not adequately model some essential features of human disease; therefore, they are often unable to pave the way to the development of drugs effective in human patients. The rise of new technological tools and models in life science, and the increasing need for multidisciplinary approaches have encouraged many interdisciplinary research initiatives. With considerable funds being invested in biomedical research, it is becoming pivotal to define and apply indicators to monitor the contribution to innovation and impact of funded research.
2. Introduction
Noncommunicable diseases (NCDs) are becoming increasingly prevalent in Western countries, and globally account for more than 80% of total premature deaths [1]. NCDs are generally the result of a combination of genetic, physiological, environmental, and lifestyle factors, with both young and middle-aged people affected. The economic consequences are also important, with lowering of earnings, employment rates, and productivity; enormous societal impacts; and health care costs. The negative effects on the gross domestic product (GDP) for EU economies are estimated to be in the order of 115 billion euros, corresponding to 0.8% of GDP annually [2].
Among NCDs, dementia, including Alzheimer’s disease (AD), affects nearly 50 million people worldwide [3]. According to the 2016 World Health Organization (WHO) statistics, AD and other dementias represent the fifth leading cause of death globally [4], and since 2015, AD has become the first leading cause of death in the UK [5]. In 2015, the number of people living with dementia in Europe was estimated to be 10.5 million (with AD accounting for 60% to 80% of dementia cases) [6].
Cancer is the second leading cause of death globally, with nearly one in six deaths caused by cancer [1]. According to the 2018 world cancer statistics [7], about 18 million new cases were diagnosed in 2018 with cancer (9.5 million men and 8.5 million women), with lung cancer, breast cancer (BC), colorectal cancer, and prostate cancer (PC) representing the four most common cancers globally (Table 1).
Table 1. The four most common cancers worldwide (modified from the 2018 world cancer statistics [7]).
| Ranking | Cancer Type | New Cases Diagnosed in 2018 (both Sexes) | % of All Cancers (Excluding Non-Melanoma Skin Cancer) |
|---|---|---|---|
| 1 | Lung | 2,093,876 | 12.3 |
| 2 | Breast | 2,088,849 | 12.3 |
| 3 | Colorectal | 1,800,977 | 10.6 |
| 4 | Prostate | 1,276,106 | 7.5 |
In Europe (EU-28), in 2016, 97,000 people died from BC, making up around 7% of all deaths from cancer, and among women, BC accounted for 15.6% of all cancer deaths [8]. With regard to PC, 76,900 men died from PC in the EU-28 in 2016, which corresponds to 5.6% of all deaths from cancer [8].
Animal models of AD, BC and PC have been and are still largely used in some of those projects aiming to recapitulate human disease features, understand the underpinning molecular and cellular mechanisms, often with the goal of identifying new druggable targets and ultimately developing new drugs. Transgenic (Tg) animals (mainly rodents) are generally purported to be reliable proxies of human diseases [9], and ‘humanized’ animals accounting for more than one engineered genetic modification are deemed to be more suitable to study complex multigene disorders [10]. However, as a matter of fact, animal models typically do not develop the diseases as they physiologically occur in humans, and the successful translation of novel research on preclinical animal models to the clinic often remains a significant barrier [11][12][13], as it has been discussed mainly in the context of AD [14][15][16], but also with regard to BC [17][18] and PC [19][20]. Flaws in animal experimentation design; inappropriate target selectivity; neglecting efficacy, pharmacodynamic, and pharmacokinetic properties of new compounds, along with inappropriate selection of clinical trial participants are among the possible reasons behind the clinical failure rate in drug development, which still remains very high, with an overall likelihood of approval from phase I of about 9.6% [21]. Notably, 97% of drugs that are tested in clinical trials in oncology never advance to receive regulatory approval [22], with a lack of efficacy and (off-target) toxicity representing the most common causes of trial failure. With regard to AD, the failure rate of drug development reaches 99% [23], and zero disease-modifying therapies have been developed so far [24]. One of the main reasons of these failures is linked to the current drug development pipeline, which still strongly relies on animal models at the preclinical stage [25].
In an effort to develop and optimize more human relevant models and increase predictive capacity, a wide range of non-animal approaches have been developed in recent years, spanning from patient-derived cells, such as induced pluripotent stem cells (iPSCs) [26][27][28], three-dimensional (3-D) tumor spheroids [29], complex microfluidics organ-on-chip technologies [25][30], to next-generation sequencing and omics technologies, integrated computer modelling, systems biology, and imaging techniques [31][32]. These models and techniques, along with data derived from clinical and observational studies, can already be used in an integrated manner to gather insights into disease molecular and cellular mechanisms, discover new biomarkers, and design novel therapeutic and preventive strategies.
The rise of new technological tools and models in life science, and the increasing need for multidisciplinary approaches, have encouraged many research initiatives and the launch of numerous research projects funded by the European Commission (EC), in particular under Framework Programme 7 (FP7: 2007–2013) and Horizon 2020 (H2020: 2014–2020), to further develop such innovative approaches [33][34][35]. During the same period, research on NCDs, and in particular on AD, BC, and PC, has been substantially funded by the EC (Figure 1, Figure 2 and Figure 3). This research effort has contributed to numerous flagships of health research, such as the Innovative Medicines Initiative [36], which has globally aided in increasing our understanding of the mechanisms underlying disease etiopathogenesis and consolidation, with a large portfolio of different activities, including basic understanding, diagnostics, drug development, drug repurposing, clinical trials, etc. Nowadays, it is becoming pivotal to define and apply indicators suitable to monitor the contribution to innovation and the impact of funded research. In the next sections, we highlight some of the issues hampering the translation of biomedical research results, specifically in the field of AD, BC, and PC. These diseases were selected as representative case studies for NCDs for several reasons: (i) Their high prevalence; (ii) the high number of animals used, as indicated by the most recent EU statistics on the number of animals used for scientific purposes [37]; (iii) the important research investments; and (iv) the high level of translational failure in drug development.
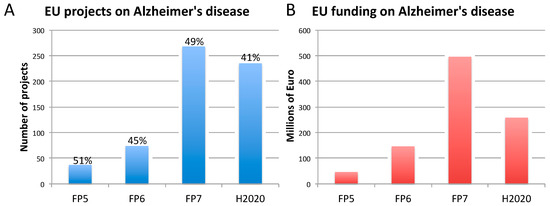
Figure 1. Number of EU-funded projects (A) and overall funding (in millions of euro) (B) allotted on Alzheimer’s disease-related research across Framework Programme (FP) 5, FP6, FP7 and Horizon 2020 (H2020). The percentages indicate the proportion of projects within a Framework Programme involving also the use of non-human animal models. Data were obtained from CORDIS [128], considering objective, reporting, and results sections, to assess whether non-human animal models were accounted for (H2020 data, as of 10 April 2020). As H2020 was still ongoing at the time of this analysis, H2020 data are not complete.
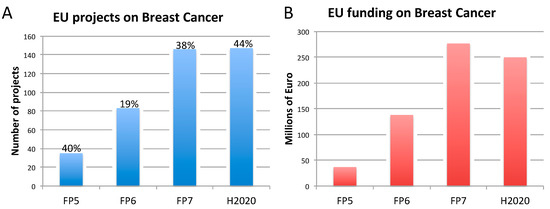
Figure 2. Number of EU-funded projects (A) and overall funding (in millions of euro) (B) allotted on breast cancer-related research across Framework Programme (FP) 5, FP6, FP7 and Horizon 2020 (H2020). The percentages indicate the proportion of projects within a Framework Programme involving also the use of non-human animal models. Data were obtained from CORDIS [128], considering objective, reporting, and results sections, to assess whether non-human animal models were accounted for. FP7 and H2020 data were obtained using the EC CORDA database (H2020 data, as of May 15, 2019). As H2020 was still ongoing at the time of this analysis, H2020 data are not complete.
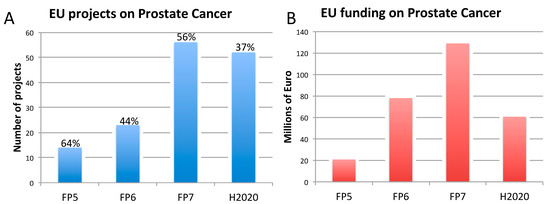
Figure 3. Number of EU-funded projects (A) and overall funding (in millions of euro) (A) allotted on prostate cancer-related research across Framework Programme (FP) 5, FP6, FP7 and Horizon 2020 (H2020). The percentages indicate the proportion of projects within a Framework Programme involving also the use of non-human animal models. Data were obtained from CORDIS [128], considering objective, reporting, and results sections, to assess whether non-human animal models were accounted for. FP7 and H2020 data were obtained using the EC CORDA database (H2020 data, as of 15 May 2019). As H2020 was still ongoing at the time of this analysis, H2020 data are not complete.
3. Three Biomedical Research Areas Characterized by a High Rate of Translational Failure: Alzheimer’s Disease, Breast Cancer, and Prostate Cancer
3.1. Alzheimer’s Disease
According to the WHO, the proportion of the world’s population over 60 years of age will nearly double from 12% to 22% between 2015 and 2050 [38]. Each year, 9.9 million new cases of dementia are registered worldwide, which corresponds to nearly one new case every 3.2 s [39]. Today, dementia affects nearly 50 million people globally, with significant public health costs [40]. Among dementias, AD accounts for about 60–80% of total cases [3].
The most common form of AD (95–99% of all AD cases) is the late-onset type (LOAD), often sporadic, generally occurring after 65 years of age, while about 1–5% of AD cases are early onset, including familial (genetically driven) (FAD) and sporadic forms. Early onset FAD generally appears before 60–65 years of age, and often before 55 years of age. Three genes are known to cause FAD: Amyloid precursor protein (APP), presenilin-1 (PS1), and presenilin-2 (PS2), with PS1 mutations accounting for most of early onset FAD [41]. Additionally, the ε4 allele of apolipoprotein E (APOE4) is a known genetic risk factor, which increases the likelihood to develop LOAD, with homozygous APOE4 carriers being nearly 15 times more likely to develop the disease [42].
All AD forms are generally characterized by the accumulation of amyloid beta (Aβ) plaques and hyperphosphorylation of tau, the major microtubule-associated protein, causing neurofibrillary tangle (NFT) formation. The disease develops with progressive cerebral atrophy, cognitive function decline, and ultimately death [43][44]. However, the role of some etiological mechanisms and contributing factors implicated in the onset and the consolidation of AD, both FAD and LOAD, still remain not fully elucidated.
Concerning drug development, despite very encouraging results obtained in preclinical animal models, showing a reduction of NFTs, APP, or Aβ plaques, often accompanied with significant improvement of spatial learning and memory, therapeutic approaches based on the ‘amyloid cascade’ hypothesis or designed to target tau proteins have generally failed to provide beneficial effects in AD patients [45][46]. One possible explanation for the failure of clinical trials is the time when drugs are being given, considering that Aβ plaques can form decades before the appearance of first cognitive symptoms [47]. Further complicating the relationship between Aβ plaques and AD, studies on post-mortem brains have reported zero or minimal levels of brain Aβ plaques in about 14–21% of clinically diagnosed patients [48][49][50].
To date, only five drugs, i.e., memantine (an antagonist of the N-Methyl-D-Aspartate-receptor subtype of glutamate receptor), three acetylcholinesterase (AChE) inhibitors (donepezil, galantamine, and rivastigmine), and a combination of donepezil with memantine, have been officially approved for the treatment of AD [15][51]. While these drugs can help temporarily improve cognitive and behavioral symptoms in AD patients, they have no effects on the long-term prognosis, they are not effective in all patients, and those patients who will respond to treatments cannot be identified early, as reported for rivastigmine, galantamine, and donepezil [52]. As indicated in a systematic review and meta-analysis study on the effects of donepezil in 8257 participants with mild, moderate, or severe AD, the quality of the evidence of donepezil’s beneficial effects on cognitive function and daily living activities, after 12 or 24 weeks of treatment, has been rated as moderate, generally due to study limitations [53].
In the last 15 years, there have been no approved disease-modifying treatments for AD. In a 2019 analysis of the drug development pipeline for AD (conducted by using the clinicaltrials.gov database), 132 agents were in clinical trials for the treatment of AD (28 agents were in 42 phase III trials). Of the several drugs that have completed clinical trial evaluation since the 2018 pipeline analysis conducted by the same group, none of them have shown a drug–placebo difference. Those clinical trials have been terminated, often upon futility analysis (which probes the ability of a clinical trial to achieve its objectives [54]), such as for crenezumab, aducanumab, verubecestat, lanabecestat, intranasal insulin, pioglitazone, AZD0530, and ITI-007; others have been terminated due to the appearance of adverse effects (e.g., atabecestat). Of the 17 disease-modifying treatments that were in phase III according to the authors’ previous 2018 review, 8 were terminated (as of 12 February 2019) [55].
The reasons underlying failure in AD drug development may be associated to some major issues, as highlighted by Mullane and Williams in their recent perspective article [15]. In particular, during preclinical drug assessment, some relevant aspects, such as target selectivity, efficacy, and the pharmacodynamic and pharmacokinetic properties of new compounds [56][57][58][59], are sometimes neglected or not taken into sufficient account, making compounds proceed to later phases of clinical trials even in the absence of clear evidence of drug efficacy [60][61]. For instance, this has happened for compounds targeting amyloid deposition, which, despite promising effects in preclinical animal models, have been shown to be ineffective in clinical trials [14][62].
Along with this, there has been the tendency to conceive AD exclusively as an amyloid- and NFT-related disease, and therefore to design drugs aimed at targeting these pathological features, not reckoning the multifactorial nature of this disease, encompassing metabolism, immune system, and inflammation, along with environmental and lifestyle-related factors [63].
Additionally, it is generally assumed that LOAD, the most prevalent AD form, and the less frequent early onset FAD are essentially the same type of disease. As both FAD and LOAD are characterized by the same pathological and behavioral traits [64], the development of drugs or new druggable targets suitable for LOAD have been studied in Tg mouse models that overexpress human genes associated with FAD. However, these models have been shown neither to recapitulate the genetics, nor the onset and progression of LOAD as observed in patients [14][65][66]. Tg and double-Tg animals develop Aβ plaques and associated brain inflammation, undergoing cognitive and behavioral deficits. These models do not produce NFTs, which can be observed along with cognitive deficits in animals engineered to express the mutated tau protein. The triple 3xTg mice (mean lifespan of 12–18 months) express mutated human APP, PSEN1, and tau protein and can generate both Aβ plaques and NFTs, producing also gliosis, synaptic deficits, and memory impairment [67].
Of the 180 available mouse models of AD (Table S1) [68]), very few are representative of LOAD [69]. One of them could be the senescence-accelerated mouse prone 8 (SAMP8), with a mean lifespan of 9.7 months, and characterized by a spontaneous accelerated aging phenotype, which is considered more suitable to study brain ageing and LOAD [70]. SAMP8 mice develop Aβ plaques, NFTs, and hyperphosphorylated tau, exhibiting spongiosis, gliosis, forebrain cholinergic impairments, and dendritic spine loss [71]. Additionally, variant strains of the SAMP8 model (e.g., SAMP8-APP, SAMP8-PS1, and SAMP8-APP/PS1 models) [72], and the more recently developed App KO/APOE4/Trem2*R47 mouse model [73], represent additional alternate models considered suitable to study LOAD.
Currently available treatments have shown beneficial effects in SAMP8 mice. For instance, memantine was found to improve spatial learning and memory and reduce both hippocampal CA1 NFTs and APP levels when administered to SAMP8 mice; additive effects were observed by combining memantine with environmental enrichment [74]. Along the same line, donepezil was found to improve spatial learning and memory ability, increase cerebral glucose metabolism, and reduce Aβ levels in the cortex of SAMP8 mice. When combined with manual acupuncture, additive beneficial effects were observed [75]. However, despite the promising results obtained in preclinical trials, clinical trials have not proven significant beneficial effects, especially in the long term. In particular, memantine treatment has shown unclear positive effects in patients, slowing the process of cognitive loss at most [76]. Combination therapies are considered more promising than individual treatments in slowing cognitive decline; for example, administration of memantine, in combination with AChE inhibitors (e.g., donepezil or galantamine) was shown to provide some behavioral benefits in patients affected by moderate to severe AD [77][78].
Despite the extensive characterization of the SAMP8 and the SAMP8 murine variants, the genes responsible for the accelerated senescence and the exhibited pathological features are almost unknown. Moreover, Aβ plaque formation and cognitive abnormalities in these mice appear to be significantly different from human AD [79].
Additionally, Tg animals expressing FAD genetic variants (Table S1), despite showing amyloids, NFTs, gliosis, and synaptic alterations, often do not undergo significant neuronal loss, and the amyloid peptides they generate appear to be different from those identified in the human brain [80]. Another relevant aspect is that animal models of AD generally do not reflect the pathology as observed in humans [16][81], and do not develop the typical co-morbidities observed in AD patients, such as metabolic syndrome, cardiovascular disease, inflammation, and immunological disorders [15]. Notably, different murine strains show remarkably different lifespans, often premature mortality, along with sex differences in the expected lifespan, as summarized by Rae and Brown [82]. Although efforts to translate lifespan developmental stages in mice to the equivalent stages for humans have been made considering chronological ages [83], these comparisons have been based on the C57 black 6 (C57BL/6) mouse, which is one of the murine strains with the longest lifespan [84]. These differences make direct comparison between preclinical studies using different murine strains quite hard, and the translation of mouse data to human clinical studies questionable. Altogether, these animal–human discrepancies have contributed to making basic science research outcomes poorly applicable to human AD [50].
3.2. Breast Cancer
Nowadays, the average 5-year survival rate for BC is 91%, compared to the 53% of those diagnosed in the 1970s [85][86]. Despite this positive trend, BC still remains the most commonly occurring cancer in women, with over two million new cases and more than half a million of diagnosed women succumbing every year [87].
To date, there are 34 drugs approved for BC treatment [88][89], a relatively high number, compared to other solid tumors. However, only 5% of molecules that show anticancer activity in preclinical studies are approved upon demonstration of sufficient efficacy in phase III clinical trials [90]. This trend is extremely prevalent especially for vascular endothelial growth factor (VEGF) inhibitors (e.g., Bevacizumab), which are also used for BC treatment [91]. As reported for AD [56][57][58][59][60][61], incorrect identification of drug targets, improper proposed mechanism of action, drug toxicity, drug resistance, and weak genetic evidence are among the possible failure reasons [22].
For example, although hormone therapies (e.g., tamoxifen, inhibitors of aromatase enzymes involved in estrogens synthesis) and Herceptin (trastuzumab) have improved clinical outcome of poor prognosis for estrogen receptor (ER)-positive and human epidermal growth factor receptor 2 (HER2)-positive cancers, respectively, many patients develop progressive disease [92][93][94]. This suggests that, despite drugs successfully passing preclinical and clinical phases, reaching marketing approval, they may still prove ineffective in the long term. Assessment of drug efficacy in the long term would not be possible in animal models, considering the way preclinical animal experimentation is generally designed (i.e., based on animals, such as mice, with limited lifespan, and treated for relatively short periods of time), which does not allow the prediction of possible drug resistance and disease progression.
Understanding de novo or acquired resistance to these drugs is one of the biggest challenges in the identification of new effective therapeutic agents. As commented by Moissenko et al. in their perspective article [95], both intracellular (e.g., drug metabolism and efflux, target modulations, lesion restoration) and extracellular mechanisms (e.g., crosstalk between tumor cells and environmental factors) may be responsible for drug resistance in BC. Although several mechanisms underlying tumor cell resistance to conventional cytotoxic compounds have been elucidated, more research is warranted to elucidate how multidrug resistance occurs in patients with advanced BC [95].
The story of iniparib (a poly (ADP-ribose) polymerase 1 (PARP1) inhibitor) teaches us the importance of comparators and specialized populations’ selection in clinical trials. Combined with chemotherapy, iniparib increased the rate of response to 52% (from 32% in the chemotherapy-alone group), suggesting that it may overcome the intrinsic drug resistance of some triple-negative BCs [96]. However, these promising data were not confirmed in the phase III trial. The fact that patients in the control group of this trial were permitted to crossover to iniparib has been implicated as potentially biasing the overall survival results. Further studies suggest that the actual mode of action of iniparib is different from what was originally expected and that its beneficial effects are largely restricted to Breast Related Cancer Antigen (BRCA)-mutation carriers [97]. Iniparib is a stark example of how incorrect interpretation of preclinical data in animals, along with poor clinical trial design, may skew results interpretation and decision-making, leading to late-stage trial failure. As highlighted by Mateo et al. [98], collected preclinical data could not elucidate the mechanism of action of iniparib before the initiation of clinical trials, and phase I trials did not prove the mechanism of action of this drug. Additionally, inappropriate selection of patients and the lack of implementation and validation of predictive biomarkers can further contribute to clinical failure. These are just some of the critical factors to be carefully considered during the development of anticancer drugs, in order to minimize failures in future late-stage clinical trials.
The greatest challenge to defeat BC is linked to the high tumor heterogeneity. Indeed, it is considered a combination of heterogeneous-related diseases, each with its specific histopathological, genomic and proteomic characteristics [99]. Heterogeneity seems to be more frequent within, not across, different BC subtypes: Only a small part of the mutations found in the primary tumor are detectable in the metastatic lesion, indicating significant genetic evolution occurring during the metastatic process [99].
Clinically, BC is categorized into three basic therapeutic groups: (i) The ER positive, (ii) the HER2 positive, and (iii) the triple negative (ER/PR/HER2 negative, where PR stands for progesterone receptor) for which no targeted therapy is currently available [100], besides chemotherapy.
Although advances in sequencing analysis have enabled the identification of many mutations, their role in disease progression is not fully understood. The Cancer Genome Atlas (TCGA) [101] has molecularly characterized over 20,000 primary cancers, and matched normal samples spanning 33 cancer types. These data, in combination with validated in vitro or in silico studies, could provide a powerful tool to explore important genomic trends or individual genes involved, possibly enabling personalized medicine approaches.
The key to success in drug development and clinical treatments is the availability of reliable preclinical models that accurately recapitulate the relevant clinical features of the disease, and therefore allow reliable screening of anticancer agents with robust clinical correlation. These models should be able to catch the whole complexity and heterogeneity of BC. Beside cell cultures, a large number of animal models are available, reflecting different types and stages of the disease. It is important to select the correct model depending on the research question(s): Characterization of the different stages of the disease, the role of the immune system and of the microenvironment, and the metastatic disease and the pathway(s) responsible for drug resistance. Advantages and pitfalls of commonly used BC models are commented in [102][103] and briefly described below.
BC cell lines have been largely used since the 1970s as in vitro models for drug discovery [104]. Cells are cultivated in an artificial environment that usually select specific populations and induce changes to facilitate cell adaption to the artificial culture environment (i.e., plastic). Continuous cell passaging might cause clonal selection and consequent loss of heterogeneity following disruption of the original tumor structure and microenvironment. Gisselsson et al. [105] comprehensively reported about the possible causes of clonal selection, specifically identifying (i) alterations in telomere function occurring over prolonged in vitro culture, and (ii) population (or genetic) bottlenecks (i.e., a significant decrease in the size of a biologically reproductive population that could be caused by factor(s) limiting the number of cells allowed to proliferate) as frequently neglected phenomena that may cause alterations in (cancer) cell line genotypes even after few passages in vitro. Compared to patient tumors, BC cell lines show a higher mutation frequency, which, over many in vitro passages, may originate a cell different from the original one. Mutations and chromosomal instability can trigger genomic heterogeneity, altering the transcriptional profile and drug response of cell lines. As a consequence, a candidate drug might be effective on the selected cellular population but fail in the clinical trial. This could explain the contradictory results observed by comparing data obtained using the same cell lines in different non-validated studies [106]. Additionally, cancer cells cultivated in vitro do not include key factors, such as stromal cells, immune and inflammatory cell infiltration, and vascularity, whose finely regulated interplay is responsible for tumor growth and metastasis formation.
Cell line-derived xenograft (CDX) models are generated by transplanting immortalized human cancer cell lines into immunocompromised mice. The choice of the transplantation site, i.e., ectopic (via subcutaneous injection) or orthotopic (in the mouse mammary gland), is critical, as each site has its own microenvironment and vasculature affecting the tumor growth rate and drug delivery [107][108]. However, CDX models lack the broad molecular transformation events (intratumoral heterogeneity) that occur in human tumors and the organotypic tumor microenvironment; therefore, they cannot recapitulate what is observed in patients with particular respect to drug response, and have been shown to poorly predict clinically effective therapies [99]. CDX models very rarely develop spontaneous metastases, making their use to study BC metastasis questionable [102]. Moreover, the cell lines used to generate CDX are generally obtained from highly aggressive tumors or fluids that have been drained from lung metastasis (e.g., MDA-MB-231 cells), which make these models less suitable to studying early events in the evolution of the primary tumor [102]. Furthermore, CDX models seem to be more responsive to antiproliferative agents than primary tumor [107].
Despite these limitations, CDX models are still widely applied at early stages of in vivo studies both in academia and industry for their user-friendly technique and high reproducibility, and several CDX mouse models have been made available by animal models’ providers (e.g., [109][110]).
Patient derived xenograft models (PDX) are obtained by surgical implantation of patient-derived tumor explant into an immunocompromised mouse. Although this method does not require preculture of patient cells, it requires fresh patient material and operator expertise, is invasive, and is rather expensive [99]. Months are needed to establish the engraftment and develop preclinical research samples, a period often too long for clinician decision-making (weeks). Compared to CDX models, PDX models retain the genetic intra/intertumor heterogeneity of the original tumor. They reflect more accurately the human situation due to contextual incorporation of human stroma and associated vasculature and tumor-associated immune cells; although, after three to five passages following engraftment, the replacement with murine stroma represents an issue [111][112]. PDX models seem to recapitulate human tumor angiogenesis [113]; therefore, they are often used for evaluating antiangiogenic therapies. The predictive power of PDX models has led to co-clinical trials, where patients and PDX models implanted with the patient tumor are treated simultaneously [113]. Immunocompromised hosts, such as severely compromised immune-deficient (SCID) mice, non-obese diabetic (NOD)–SCID mice, athymic nude mice, recombination-activating gene 2 (Rag2)-knockout mice, and the NOD/SCID/IL2Rγc−/− mice, are frequently used to generate the PDX model of BC [114], as they allow tumor engraftment. However, as these models lack immune system cells, they are not suitable for the preclinical testing of immunotherapies [99]. Murine strains with a human immune system (“humanized mice”), generated by engrafting different types of human leukocytes and purified human hematopoietic stem cells (CD34+) obtained from bone marrow, umbilical cord blood cells, fetal livers, or thymus tissues, are currently regarded as suitable models for immunotherapy efficacy testing [114]. However, co-engraftment with human immune cells still presents some limitations (e.g., xenogeneic graft-versus-host responses) [115], and introduces an extra layer of complexity and costs associated with their generation and maintenance [116][117]. In PDX models, aggressive tumor subtypes are usually favored and these forms might not respond to therapy as the less aggressive forms. As they are generated at the point of BC surgery, only after a period of at least five years it would be possible to compare the PDX model’s capacity to form metastases to what is observed in the patient. The average mouse lifespan is about 2 years, and, in the case of the commonly used NOD SCID murine strain, the lifespan is approximately 30 weeks, which is often due to the spontaneous development of thymic lymphomas. Therefore, these models are not deemed suitable for long-term xenotransplantation studies [118].
At a clinical level, metastatic lesions are often vastly different from their primary tumor counterparts at both molecular and histological levels. Lungs and lymph nodes metastases are commonly observed in PDX models, while the high frequency of brain and bone metastases observed in patients (approximately 70%) are rarely observed [102][119].
Genetically engineered mouse models (GEMMs) are generally used to address early events in the tumor process, as these models spontaneously exhibit tumor initiation driven by oncogenes within the correct microenvironment (e.g., MMTV-PyMT). However, the lineage/expression domains of the regulatory sequences used to induce transgene expression are not well defined, and GEMM oncogenes may not necessarily be representatives of those observed in human tumors [102]. Specific Tg animals have been developed in an effort to emulate more closely the genetics of human BC, accounting for the temporal and spatial activation of specific oncogenes and the deletion of tumor suppressors targeted to the murine mammary gland, such as the Cre/loxP conditional BLG-Cre;Brca1F22−24;p53KO model [120]. Mouse and rat models of BC have been customized using the nuclease-based system Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR)/CRISPR associated protein 9 (CRISPR/Cas9), which can target any gene within a eukaryotic genome, and have been made available by different animal vendors. Comprehensive lists of BC GEMMs along with their characteristics have been described in several review articles [121][122][123][124][125].
Considering the very high level of tumor heterogeneity observed in BC, with specific histopathological, genomic, and proteomic features, and the genetic evolution frequently observed during metastasis [99], the use of animal models (in particular, CDX and GEMM models) to test new drugs may not be the best methodological approach to account for this complexity and understand BC biology and evolution as it occurs in humans.
None of the above-mentioned models seem to successfully recapitulate the metastasis process in BC. Although the 5-year survival rate for metastatic BC is 22% [86], and there is reasonable evidence that some patients with metastatic BC can be cured [126], less than 5% of global funding for BC research is dedicated toward understanding metastatic BC, or finding solutions to extend the lives of metastatic BC patients [127].
By the time a woman is diagnosed with metastatic disease, her original biopsy tissue may no longer be available. In addition, it is often hard to get a sample of a metastatic tumor, which may be buried inside the brain or in an anatomical area that is difficult to access safely. Current metastatic therapies are expensive, often toxic, and subject to the eventual development of drug resistance. Besides drug treatment, earlier detection of metastatic disease is of paramount importance in the prognosis of the patient. Several EU projects have been funded for the development of advanced technologies, such as positron emission tomography/computed tomography scans, or to improve imaging resolution [128]. Besides metastasis detection, these imaging technologies could also help in the early detection of tumor, saving many lives [129]. The possibility of measuring circulating tumor DNA, although not very informative in terms of time for metastatic disease development compared to cancer antigen 27-29 (CA 27-29) detection, might allow effective therapy development for a (micro/early) metastatic lesion [130].
3.3. Prostate Cancer
Prostate cancer is the second most frequent diagnosed cancer in men and the fourth most commonly occurring cancer overall (Table 1). Annually, there are more than 1.2 million new cases of PC and over 350,000 deaths, with higher incidence rates and prevalence in developed countries [131]. PC incidence and mortality rates are strongly related to age, with the highest incidence seen in elderly men (>65 years of age). The etiology of PC remains largely unknown compared to other common cancers; however, risk factors include advanced age, ethnicity, genetic factors, and family history [132].
Differences in the incidence rates worldwide are likely due to the use of different diagnostic testing and advanced screening in different regions of the world. Despite advances made in the field, PC is still a major cause of morbidity and mortality, and the overall high global incidence rate stresses the need to strengthen prevention measures, diagnostic tools, and novel therapies to reduce the public health impact of this disease.
The field of PC research continues to be hindered by the lack of human-relevant preclinical models to study disease development and progression and to further knowledge of effective prevention and therapeutic strategies [133]. PC is a highly heterogeneous and complex disease, and the research has been obstructed by the use of imperfect animal models that do not accurately recapitulate the illness, are costly, take considerable time to develop, and fail to mimic the multifaceted aspects of the condition. The most widely used animals in human PC research include genetically engineered mice, xenograft mice, Tg rats, and dogs [134]. Nevertheless, all these models have limitations that present serious challenges to preclinical drug development and biomedical research. Differences in size and physiology, as well as variations in the homology of targets between animals and humans, have led to translational limitations. For instance, mice used in PC research rarely develop tumors that metastasize, making it almost impossible to study the terminal lethal events in cancer progression [135]. Other than humans, dogs are the only other species that develop benign prostatic hyperplasia and sporadic PC; however, male dogs experience a very low incidence of spontaneous PC and are generally androgen independent and lack a functional androgen receptor (AR), unlike PC in humans [134].
When taking a closer look at the most extensively used animal in PC research, the mouse, there are several disadvantages that may explain why so few drugs that work in mice do not work in humans. With regard to prostate gene expression, while both mouse and human prostate cells respond to androgen stimulation and signaling [136], the expression of certain human androgen-responsive genes (e.g., prostate-specific antigen (PSA) and prostate-specific membrane antigen (PSMA)) is not present in mice [137][138][139]. When analyzing the prostates of mice and men, they significantly differ anatomically. Human prostates consist of a single lobe and three zones (central, peripheral, and transitional) surrounding the urethra at the base of the bladder. In mice, the prostate is comprised of four lobes located circumferentially around the urethra. Testosterone levels also fluctuate between the two species, and significant interspecies differences have been described with regard to the serum protein-binding affinity for androgens, regulation and function of hepatic steroid metabolizing enzymes, and testosterone biosynthesis and metabolism [140]. In mice, the total and free plasma testosterone levels may vary drastically between individual mice and between genetic backgrounds [138]. Additionally, the histopathology and timeframe of PC development is different in mice [141].
Despite the worldwide research endeavors, few findings have influenced the clinical management of the disease [19]. When it comes to drug discovery and development, CDX and PDX mouse models, where human PC cells are transplanted into immune-deficient mice, are the most often used. However, this is problematic for many reasons; as it has been already commented in Section 2.2 (on BC), research has shown that the immune system plays an important role in cancer progression and eradication, yet the immune system cannot be studied in immune-deficient mice. Another issue with using CDX mice is that they fail to reproduce the diverse heterogeneity observed in humans, partially due to the increased homogeneity of established cell lines after long-term in vitro culturing and the relative lack of cell lines in PC [138]. The use of immune-deficient xenotransplanted mice for the development of hormone ablation therapies presents significant limitations, as these animals present inadequate (low) levels of testosterone, not reflecting those found in normally ageing men [142].
Several GEMMs of PC have been developed, by modulating the expression of specific oncogenes or tumor suppressors, growth factors and their receptors, steroid hormone receptors, or regulators of cell cycle and apoptosis, as summarized in [143][144]. An example is provided by the PB-Cre4xPTENloxp/loxp GEMM model of PC, which is considered suitable to study prostate adenocarcinoma development, tumor progression, and metastasis [144]. Similarly, the Tg adenocarcinoma of the mouse prostate (TRAMP) model has been shown to recapitulate both the preneoplastic and metastatic stages of PC [145]. However, these models also present intrinsic limitations [146]. In particular, the TRAMP model develops primarily neuroendocrine tumors, is based on an androgen-dependent promoter, rarely undergoes bone metastasis, and exhibits relatively short kinetics opposite to the typically slow development of PC in humans [147]. Along the same line, the PTEN conditional model described above has been shown to develop senescence, which limits cancer progression, and does not develop metastases [147]. These differences in how PC spontaneously develops and evolves in humans and how it is artificially recreated in animals can possibly explain why drugs that are effective in mice (and other animals) are very often not successful in humans. It comes as no surprise that of the hundreds of drugs tested in mice to reduce tumor volume, only a few have translated into a drug effective for treating PC in humans [138]. As commented in Section 2.2., only about 5% of anticancer therapies tested in animals demonstrate sufficient efficacy in phase III clinical trials and are ultimately approved for clinical use [90]. The cost of these failures is estimated in the range of hundreds of millions of dollars per drug [148].
One drug, Orteronel (also known as TAK-700), is a hormonal therapy that was tested for the treatment of PC. Orteronel was designed to inhibit the 17,20 lyase activity of the enzyme CYP17A1, which is important for androgen synthesis in the testes, adrenal glands, and PC cells. Preclinical studies in animals provided the rationale for testing Orteronel in PC patients. In particular, Orteronel treatment caused a significant suppression of serum testosterone levels, shrinking several androgen-dependent organs in uncastrated rats [149]. Moreover, administration of Orteronel (twice daily) in intact cynomolgus monkeys induced a reduction of serum dehydroepiandrosterone sulfate and testosterone levels vs. vehicle control, and in castrated monkeys, such effects were even greater and persisted throughout the treatment period [150][151]. Despite these promising effects in animals, during phase II clinical trials, 34% of participants experienced serious adverse effects, and 96% experienced other adverse effects as a result of the treatment [152]. In a larger phase III study with 732 participants receiving Orteronel intervention, 52% were affected by serious adverse events and 96% experienced other adverse effects [153]. Ultimately, the drug did not move forward since it did not extend the overall survival of the patients and caused significant adverse effects.
Another drug that initially showed promise in preclinical studies but failed during clinical trials is AZD3514. AZD3514 is an oral drug that targets AR function, with a novel mechanism of action that can result in downregulation of AR protein. In several in vivo studies using rats, this drug significantly inhibited testosterone-induced growth of the prostate and seminal vesicles and also reduced tumor AR [154]. This drug went on to two phase I clinical trials, where only 13% of patients treated with AZD3514 had a greater than 50% decline in PSA and 17% of patients had decreased clinical indicators of soft tissue disease [155]. Despite several PSA responses observed using this therapy on animals, only marginal outcomes were detected at the clinical level and the development of AZD3514 has been discontinued due to toxicity concerns and significant adverse effects [156].
There are currently 15 approved drugs on the market for various treatment options, yet PC was still the cause of 358,989 deaths in 2018 [131][157]. Improving PC research to the point at which it translates to reproducible and effective success in the clinic is a particularly difficult challenge. Considering the heterogeneity and complexity of the disease, more resources should focus on the use of human biology-based research models complementary (or alternative) to animals, to develop promising therapeutics for all forms of PC.
This entry is adapted from the peer-reviewed paper 10.3390/ani10071194
References
- EC. Noncommunicable Diseases—NCDs. Available online: https://ec.europa.eu/knowledge4policy/foresight/topic/shifting-health-challenges/non-communicable-diseases-ncds_en (accessed on 23 April 2020).
- OECD. Health at a Glance: Europe 2018; OECD: Paris, France, 2018.
- Alzheimer's Association. 2019 Alzheimer's disease facts and figures. Alzheimer Dement. 2019, 15, 321–387.
- WHO. The Top 10 Causes of Death. Available online: https://http://www.who.int/news-room/fact-sheets/detail/the-top-10-causes-of-death (accessed on 23 April 2020).
- Deaths Registered in England and Wales (Series DR): 2017. Available online: https://http://www.ons.gov.uk/peoplepopulationandcommunity/birthsdeathsandmarriages/deaths/bulletins/deathsregisteredinenglandandwalesseriesdr/2017 (accessed on 29 June 2020).
- Is Europe Ready for Alzheimer’s? Available online: http://www.aal-europe.eu/is-europe-ready-for-alzheimers/(accessed on 29 June 2020).
- Worldwide Cancer Data. Available online: https://http://www.wcrf.org/dietandcancer/cancer-trends/worldwide-cancer-data (accessed on 23 April 2020).
- EC. Cancer Statistics—Specific Cancers. Available online: https://ec.europa.eu/eurostat/statistics-explained/pdfscache/39738.pdf (accessed on 29 June 2020).
- Simmons, D. The use of animal models in studying genetic disease: Transgenesis and induced mutation. Nat. Educ. 2008, 1, 70.
- Labant, M. Animal Models Evolve to Satisfy Emerging Needs. Available online: https://http://www.genengnews.com/insights/animal-models-evolve-to-satisfy-emerging-needs/(accessed on 29 June 2020).
- Pound, P.; Ritskes-Hoitinga, M. Is it possible to overcome issues of external validity in preclinical animal research? Why most animal models are bound to fail. J. Transl. Med. 2018, 16, 304, doi:10.1186/s12967-018-1678-1.
- Mak, I.W.; Evaniew, N.; Ghert, M. Lost in translation: Animal models and clinical trials in cancer treatment. Am. J. Transl. Res. 2014, 6, 114–118.
- Gould, S.E.; Junttila, M.R.; De Sauvage, F.J. Translational value of mouse models in oncology drug development. Nat. Med. 2015, 21, 431–439, doi:10.1038/nm.3853.
- Drummond, E.; Wisniewski, T. Alzheimer’s disease: Experimental models and reality. Acta Neuropathol. 2017, 133, 155–175, doi:10.1007/s00401-016-1662-x.
- Mullane, K.; Williams, M. Preclinical Models of Alzheimer’s Disease: Relevance and Translational Validity. Curr. Protoc. Pharmacol. 2019, 84, e57, doi:10.1002/cpph.57.
- Cavanaugh, S.E.; Pippin, J.J.; Barnard, N.D. Animal models of Alzheimer disease: Historical pitfalls and a path forward. Altex 2014, 31, 279–302, doi:10.14573/altex.1310071.
- Manning, H.C.; Buck, J.R.; Cook, R.S. Mouse Models of Breast Cancer: Platforms for Discovering Precision Imaging Diagnostics and Future Cancer Medicine. J. Nucl. Med. Off. Publ. Soc. Nucl. Med. 2016, 57(Suppl 1), 60S-68S, doi:10.2967/jnumed.115.157917.
- Ozdemir, B.C.; Sflomos, G.; Brisken, C. The challenges of modeling hormone receptor-positive breast cancer in mice. Endocr. -Relat. Cancer 2018, 25, R319–R330, doi:10.1530/ERC-18-0063.
- Rea, D.; Del Vecchio, V.; Palma, G.; Barbieri, A.; Falco, M.; Luciano, A.; De Biase, D.; Perdona, S.; Facchini, G.; Arra, C. Mouse Models in Prostate Cancer Translational Research: From Xenograft to PDX. Biomed. Res. Int. 2016, 2016, 9750795, doi:10.1155/2016/9750795.
- Hensley, P.J.; Kyprianou, N. Modeling prostate cancer in mice: Limitations and opportunities. J. Androl. 2012, 33, 133–144, doi:10.2164/jandrol.111.013987.
- bio.org. Clinical Development Success Rates 2006–2015. Available online: https://http://www.bio.org/sites/default/files/legacy/bioorg/docs/Clinical Development Success Rates 2006-2015—BIO, Biomedtracker, Amplion 2016.pdf (accessed on 23 April 2020).
- Lin, A.; Giuliano, C.J.; Palladino, A.; John, K.M.; Abramowicz, C.; Yuan, M.L.; Sausville, E.L.; Lukow, D.A.; Liu, L.; Chait, A.R.; et al. Off-target toxicity is a common mechanism of action of cancer drugs undergoing clinical trials. Sci. Transl. Med. 2019, 11, doi:10.1126/scitranslmed.aaw8412.
- Cummings, J.L.; Morstorf, T.; Zhong, K. Alzheimer’s disease drug-development pipeline: Few candidates, frequent failures. Alzheimer’s Res. Ther. 2014, 6, 37, doi:10.1186/alzrt269.
- Cummings, J.; Reiber, C.; Kumar, P. The price of progress: Funding and financing Alzheimer’s disease drug development. Alzheimer’s Dement. 2018, 4, 330–343, doi:10.1016/j.trci.2018.04.008.
- Miccoli, B.; Braeken, D.; Li, Y.E. Brain-on-a-chip Devices for Drug Screening and Disease Modeling Applications. Curr. Pharm. Des. 2018, 24, 5419–5436, doi:10.2174/1381612825666190220161254.
- Eglen, R.M.; Reisine, T. Human iPS Cell-Derived Patient Tissues and 3D Cell Culture Part 2: Spheroids, Organoids, and Disease Modeling. Slas Technol. 2019, 24, 18–27, doi:10.1177/2472630318803275.
- Essayan-Perez, S.; Zhou, B.; Nabet, A.M.; Wernig, M.; Huang, Y.A. Modeling Alzheimer’s disease with human iPS cells: Advancements, lessons, and applications. Neurobiol. Dis. 2019, 130, 104503, doi:10.1016/j.nbd.2019.104503.
- Shirotani, K.; Matsuo, K.; Ohtsuki, S.; Masuda, T.; Asai, M.; Kutoku, Y.; Ohsawa, Y.; Sunada, Y.; Kondo, T.; Inoue, H.; et al. A simplified and sensitive method to identify Alzheimer’s disease biomarker candidates using patient-derived induced pluripotent stem cells (iPSCs). J. Biochem. 2017, 162, 391–394, doi:10.1093/jb/mvx058.
- Zanoni, M.; Pignatta, S.; Arienti, C.; Bonafe, M.; Tesei, A. Anticancer drug discovery using multicellular tumor spheroid models. Expert Opin. Drug Discov. 2019, 14, 289–301, doi:10.1080/17460441.2019.1570129.
- Lee, I.C. Cancer-on-a-chip for Drug Screening. Curr. Pharm. Des. 2018, 24, 5407–5418, doi:10.2174/1381612825666190206235233.
- Silverman, E.K.; Schmidt, H.; Anastasiadou, E.; Altucci, L.; Angelini, M.; Badimon, L.; Balligand, J.L.; Benincasa, G.; Capasso, G.; Conte, F.; et al. Molecular networks in Network Medicine: Development and applications. Wiley Interdiscip. Rev. Syst. Biol. Med. 2020, 10.1002/wsbm.1489, e1489, doi:10.1002/wsbm.1489.
- Yao, F.; Hong, X.; Li, S.; Zhang, Y.; Zhao, Q.; Du, W.; Wang, Y.; Ni, J. Urine-Based Biomarkers for Alzheimer’s Disease Identified Through Coupling Computational and Experimental Methods. J. Alzheimer’s Dis. JAD 2018, 65, 421–431, doi:10.3233/JAD-180261.
- EC. FET Open. Available online: https://ec.europa.eu/programmes/horizon2020/en/h2020-section/fet-open (accessed on 23 April 2020).
- EC. Funding & Tender Opportunities. Available online: https://ec.europa.eu/info/funding-tenders/opportunities/portal/screen/opportunities/topic-search;freeTextSearchKeyword=;typeCodes=1;statusCodes=31094501,31094502;programCode=H2020;programDivisionCode=31047826;focusAreaCode=null;crossCuttingPriorityCode=null;callCode=Default;sortQuery=openingDate;orderBy=asc;onlyTenders=false;topicListKey=topicSearchTablePageState (accessed on 23 April 2020).
- Multidisciplinary Research Projects on Personalised Medicine—Pre-/Clinical Research, Big Data and ICT, Implementation and User’s Perspective. Available online: http://www.erapermed.eu/3211-2/(accessed on 23 April 2020).
- IMI. Innovative Medicines Initiative. Available online: https://http://www.imi.europa.eu/(accessed on 23 April 2020).
- EC. 2019 Report on the Statistics on the Use of Animals for Scientific Purposes in the Member States of the European Union in 2015–2017. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?qid=1581689520921&uri=CELEX:52020DC0016 (accessed on 23 April 2020).
- WHO. Ageing and Health. Available online: https://http://www.who.int/news-room/fact-sheets/detail/ageing-and-health (accessed on 23 April 2020).
- Dementia Statistics. Available online: https://http://www.alz.co.uk/research/statistics (accessed on 23 April 2020).
- Wimo, A.; Jonsson, L.; Bond, J.; Prince, M.; Winblad, B.; Alzheimer Disease, I. The worldwide economic impact of dementia 2010. Alzheimer’s Dement. J. Alzheimer’s Assoc. 2013, 9, 1–11 e13, doi:10.1016/j.jalz.2012.11.006.
- Early Onset Famlial AD. Available online: https://http://www.alzforum.org/early-onset-familial-ad/overview/what-early-onset-familial-alzheimer-disease-efad (accessed on 23 April 2020).
- Hunsberger, H.C.; Pinky, P.D.; Smith, W.; Suppiramaniam, V.; Reed, M.N. The role of APOE4 in Alzheimer’s disease: Strategies for future therapeutic interventions. Health Psychol. Behav. Med. 2019, 3, NS20180203, doi:10.1042/NS20180203.
- Daulatzai, M.A. Quintessential risk factors: Their role in promoting cognitive dysfunction and Alzheimer’s disease. Neurochem. Res. 2012, 37, 2627–2658, doi:10.1007/s11064-012-0854-6.
- World Alzheimer Report 2018. Available online: https://http://www.alz.co.uk/research/world-report-2018 (accessed on 23 April 2020).
- PCRM. Retiring the Amyloid Cascade Hypothesis as a Cause of Alzheimer’s. Available online: https://http://www.pcrm.org/news/good-science-digest/retiring-amyloid-cascade-hypothesis-cause-alzheimers (accessed on 23 April 2020).
- Ricciarelli, R.; Fedele, E. The Amyloid Cascade Hypothesis in Alzheimer’s Disease: It’s Time to Change Our Mind. Curr. Neuropharmacol. 2017, 15, 926–935, doi:10.2174/1570159 × 15666170116143743.
- Makin, S. The amyloid hypothesis on trial. Nature 2018, 559, S4-S7, doi:10.1038/d41586-018-05719-4.
- Beach, T.G.; Monsell, S.E.; Phillips, L.E.; Kukull, W. Accuracy of the clinical diagnosis of Alzheimer disease at National Institute on Aging Alzheimer Disease Centers, 2005–2010. J. Neuropathol. Exp. Neurol. 2012, 71, 266–273, doi:10.1097/NEN.0b013e31824b211b.
- Serrano-Pozo, A.; Qian, J.; Monsell, S.E.; Blacker, D.; Gomez-Isla, T.; Betensky, R.A.; Growdon, J.H.; Johnson, K.A.; Frosch, M.P.; Sperling, R.A.; et al. Mild to moderate Alzheimer dementia with insufficient neuropathological changes. Ann. Neurol. 2014, 75, 597–601, doi:10.1002/ana.24125.
- Pippin, J.J.; Cavanaugh, S.E.; Pistollato, F. Animal Research for Alzheimer Disease: Failures of Science and Ethics. In Animal Experimentation: Working Towards a Paradigm Change; Herrmann, K., Jayne, J., Eds.; Brill: Leiden, The Netherlands, 2019; doi:10.1163/9789004391192_021.
- FDA-Approved Treatments for Alzheimer’s. Available online: https://http://www.alz.org/media/documents/fda-approved-treatments-alzheimers-ts.pdf (accessed on 23 April 2020).
- Birks, J. Cholinesterase Inhibitors for Alzheimer’s Disease. Cochrane Database Syst. Rev. 2006, doi:10.1002/14651858.CD005593.
- Birks, J.S.; Harvey, R.J. Donepezil for dementia due to Alzheimer’s disease. Cochrane Database Syst. Rev. 2018, 6, CD001190, doi:10.1002/14651858.CD001190.pub3.
- Shan, G.; Banks, S.; Miller, J.B.; Ritter, A.; Bernick, C.; Lombardo, J.; Cummings, J.L. Statistical advances in clinical trials and clinical research. Alzheimer’s Dement. 2018, 4, 366–371, doi:10.1016/j.trci.2018.04.006.
- Cummings, J.; Lee, G.; Ritter, A.; Sabbagh, M.; Zhong, K. Alzheimer’s disease drug development pipeline: 2019. Alzheimer’s Dement. 2019, 5, 272–293, doi:10.1016/j.trci.2019.05.008.
- Morgan, P.; Van Der Graaf, P.H.; Arrowsmith, J.; Feltner, D.E.; Drummond, K.S.; Wegner, C.D.; Street, S.D. Can the flow of medicines be improved? Fundamental pharmacokinetic and pharmacological principles toward improving Phase II survival. Drug Discov. Today 2012, 17, 419–424, doi:10.1016/j.drudis.2011.12.020.
- De Witte, W.E.A.; Danhof, M.; van der Graaf, P.H.; de Lange, E.C.M. The implications of target saturation for the use of drug-target residence time. Nat. Rev. Drug Discov. 2018, 18, 82–84, doi:10.1038/nrd.2018.234.
- Kleiman, R.J.; Ehlers, M.D. Data gaps limit the translational potential of preclinical research. Sci. Transl. Med. 2016, 8, 320–321, doi:10.1126/scitranslmed.aac9888.
- Karran, E.; Hardy, J. A critique of the drug discovery and phase 3 clinical programs targeting the amyloid hypothesis for Alzheimer disease. Ann. Neurol. 2014, 76, 185–205, doi:10.1002/ana.24188.
- Gold, M. Phase II clinical trials of anti-amyloid beta antibodies: When is enough, enough? Alzheimer’s Dement. 2017, 3, 402–409, doi:10.1016/j.trci.2017.04.005.
- Gray, J.A.; Fleet, D.; Winblad, B. The need for thorough phase II studies in medicines development for Alzheimer’s disease. Alzheimer’s Res. Ther. 2015, 7, 67, doi:10.1186/s13195-015-0153-y.
- Pagliarulo, N.; Gardner, J. 7 Questions on Biogen’s Revival of a Failed Alzheimer’s Drug. Available online: https://http://www.biopharmadive.com/news/biogen-alzheimers-aducanumab-revival-7-questions/565609/(accessed on 23 April 2020).
- WHO. Risk Reduction of Cognitive Decline and Dementia; WHO: Geneva, Switzerland, 2019.
- Barrett, J.E.; McGonigle, P. Rodent Models for Alzheimer’s Disease in Drug Discovery. In Drug Discovery Approaches for the Treatment of Neurodegenerative Disorders; Academic Press: San Diego, CA, USA, 2017; pp. 235-247; doi:10.1016/B978-0-12-802810-0.00012-X.
- Newman, M.; Kretzschmar, D.; Khan, I.; Chen, M.; Verdile, G.; Lardelli, M. Animal models of Alzheimer’s Disease. In Animal Models for the Study of Human Disease, 2nd ed.; Michael Conn, P., Ed. Academic Press: San Diego, CA, USA, 2017; pp. 1031–1085.
- Webster, S.J.; Bachstetter, A.D.; Nelson, P.T.; Schmitt, F.A.; Van Eldik, L.J. Using mice to model Alzheimer’s dementia: An overview of the clinical disease and the preclinical behavioral changes in 10 mouse models. Front. Genet. 2014, 5, 88, doi:10.3389/fgene.2014.00088.
- Oddo, S.; Caccamo, A.; Shepherd, J.D.; Murphy, M.P.; Golde, T.E.; Kayed, R.; Metherate, R.; Mattson, M.P.; Akbari, Y.; LaFerla, F.M. Triple-transgenic model of Alzheimer’s disease with plaques and tangles: Intracellular Abeta and synaptic dysfunction. Neuron 2003, 39, 409–421, doi:10.1016/s0896-6273(03)00434-3.
- Research Models. Alzheimer’s Disease. Available online: https://http://www.alzforum.org/research-models/alzheimers-disease (accessed on 23 April 2020).
- King, A. The search for better animal models of Alzheimer’s disease. Nature 2018, 559, S13–S15, doi:10.1038/d41586-018-05722-9.
- Pallàs, M. Senescence-Accelerated Mice P8: A Tool to Study Brain Aging and Alzheimer’s Disease in a Mouse Model. Int. Sch. Res. Not. Cell Biol. 2012, 2012, 1–12, doi:10.5402/2012/917167.
- Cheng, X.R.; Zhou, W.X.; Zhang, Y.X. The behavioral, pathological and therapeutic features of the senescence-accelerated mouse prone 8 strain as an Alzheimer’s disease animal model. Ageing Res. Rev. 2014, 13, 13–37, doi:10.1016/j.arr.2013.10.002.
- Lok, K.; Zhao, H.; Zhang, C.; He, N.; Shen, H.; Wang, Z.; Zhao, W.; Yin, M. Effects of accelerated senescence on learning and memory, locomotion and anxiety-like behavior in APP/PS1 mouse model of Alzheimer’s disease. J. Neurol. Sci. 2013, 335, 145–154, doi:10.1016/j.jns.2013.09.018.
- App KO/APOE4/Trem2*R47H. Available online: https://http://www.jax.org/strain/031722 (accessed on 23 April 2020).
- Dong, J.; Zhou, M.; Wu, X.; Du, M.; Wang, X. Memantine combined with environmental enrichment improves spatial memory and alleviates Alzheimer’s disease-like pathology in senescence-accelerated prone-8 (SAMP8) mice. J. Biomed. Res. 2012, 26, 439–447, doi:10.7555/JBR.26.20120053.
- Jiang, J.; Liu, G.; Shi, S.; Li, Y.; Li, Z. Effects of manual acupuncture combined with donepezil in a mouse model of Alzheimer’s disease. Acupunct. Med. J. Br. Med Acupunct. Soc. 2019, 37, 64–71, doi:10.1136/acupmed-2016-011310.
- Folch, J.; Busquets, O.; Ettcheto, M.; Sanchez-Lopez, E.; Castro-Torres, R.D.; Verdaguer, E.; Garcia, M.L.; Olloquequi, J.; Casadesus, G.; Beas-Zarate, C.; et al. Memantine for the Treatment of Dementia: A Review on its Current and Future Applications. J. Alzheimer’s Dis. JAD 2018, 62, 1223–1240, doi:10.3233/JAD-170672.
- Grossberg, G.T.; Manes, F.; Allegri, R.F.; Gutierrez-Robledo, L.M.; Gloger, S.; Xie, L.; Jia, X.D.; Pejovic, V.; Miller, M.L.; Perhach, J.L.; et al. The safety, tolerability, and efficacy of once-daily memantine (28 mg): A multinational, randomized, double-blind, placebo-controlled trial in patients with moderate-to-severe Alzheimer’s disease taking cholinesterase inhibitors. CNS Drugs 2013, 27, 469–478, doi:10.1007/s40263-013-0077-7.
- Howard, R.; McShane, R.; Lindesay, J.; Ritchie, C.; Baldwin, A.; Barber, R.; Burns, A.; Dening, T.; Findlay, D.; Holmes, C.; et al. Nursing home placement in the Donepezil and Memantine in Moderate to Severe Alzheimer’s Disease (DOMINO-AD) trial: Secondary and post-hoc analyses. Lancet. Neurol. 2015, 14, 1171–1181, doi:10.1016/S1474-4422(15)00258-6.
- Porquet, D.; Andres-Benito, P.; Grinan-Ferre, C.; Camins, A.; Ferrer, I.; Canudas, A.M.; Del Valle, J.; Pallas, M. Amyloid and tau pathology of familial Alzheimer’s disease APP/PS1 mouse model in a senescence phenotype background (SAMP8). Age 2015, 37, 9747, doi:10.1007/s11357-015-9747-3.
- Morris, G.P.; Clark, I.A.; Vissel, B. Questions concerning the role of amyloid-beta in the definition, aetiology and diagnosis of Alzheimer’s disease. Acta Neuropathol. 2018, 136, 663–689, doi:10.1007/s00401-018-1918-8.
- Veening-Griffioen, D.H.; Ferreira, G.S.; van Meer, P.J.K.; Boon, W.P.C.; Gispen-de Wied, C.C.; Moors, E.H.M.; Schellekens, H. Are some animal models more equal than others? A case study on the translational value of animal models of efficacy for Alzheimer’s disease. Eur. J. Pharmacol. 2019, 859, 172524, doi:10.1016/j.ejphar.2019.172524.
- Rae, E.A.; Brown, R.E. The problem of genotype and sex differences in life expectancy in transgenic AD mice. Neurosci. Biobehav. Rev. 2015, 57, 238–251, doi:10.1016/j.neubiorev.2015.09.002.
- Flurkey, K.; J.M., C.; Harrison, D.E. Mouse Models in Aging Research. In The Mouse in Biomedical Research, 2nd Ed.; Medicine, A., Ed. Academic Press: Cambridge, MA, USA, 2007; Volume III, pp. 637-672.
- Yuan, R.; Peters, L.L.; Paigen, B. Mice as a mammalian model for research on the genetics of aging. Ilar J. 2011, 52, 4–15, doi:10.1093/ilar.52.1.4.
- Breast Cancer: Statistics. Available online: https://http://www.cancer.net/cancer-types/breast-cancer/statistics (accessed on 24 April 2020).
- cancer.org. Survival Rates for Breast Cancer. Available online: http://www.cancer.org/cancer/breastcancer/detailedguide/breast-cancer-survival-by-stage (accessed on 24 April 2020).
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. Ca A Cancer J. Clin. 2018, 68, 394–424, doi:10.3322/caac.21492.
- NIH. Drugs Approved for Breast Cancer. Available online: https://http://www.cancer.gov/about-cancer/treatment/drugs/breast (accessed on 23 April 2020).
- Leo, C.P.; Leo, C.; Szucs, T.D. Breast cancer drug approvals by the US FDA from 1949 to 2018. Nat. Rev. Drug Discov. 2020, 19, 11, doi:10.1038/d41573-019-00201-w.
- Hutchinson, L.; Kirk, R. High drug attrition rates--where are we going wrong? Nat. Rev. Clin. Oncol. 2011, 8, 189–190, doi:10.1038/nrclinonc.2011.34.
- Ju, J.; Zhu, A.J.; Yuan, P. Progress in targeted therapy for breast cancer. Chronic Dis. Transl. Med. 2018, 4, 164–175, doi:10.1016/j.cdtm.2018.04.002.
- Fan, P.; Jordan, V.C. New insights into acquired endocrine resistance of breast cancer. Cancer Drug Resist. 2019, 2, 198–209, doi:10.20517/cdr.2019.13.
- Li, G.; Guo, J.; Shen, B.Q.; Yadav, D.B.; Sliwkowski, M.X.; Crocker, L.M.; Lacap, J.A.; Phillips, G.D.L. Mechanisms of Acquired Resistance to Trastuzumab Emtansine in Breast Cancer Cells. Mol. Cancer Ther. 2018, 17, 1441–1453, doi:10.1158/1535-7163.MCT-17-0296.
- Luque-Cabal, M.; Garcia-Teijido, P.; Fernandez-Perez, Y.; Sanchez-Lorenzo, L.; Palacio-Vazquez, I. Mechanisms Behind the Resistance to Trastuzumab in HER2-Amplified Breast Cancer and Strategies to Overcome It. Clin. Med. Insights Oncol. 2016, 10, 21–30, doi:10.4137/CMO.S34537.
- Moiseenko, F.; Volkov, N.; Bogdanov, A.; Dubina, M.; Moiseyenko, V. Resistance mechanisms to drug therapy in breast cancer and other solid tumors: An opinion. F1000Research 2017, 6, 288, doi:10.12688/f1000research.10992.1.
- O’Shaughnessy, J.; Osborne, C.; Pippen, J.E.; Yoffe, M.; Patt, D.; Rocha, C.; Koo, I.C.; Sherman, B.M.; Bradley, C. Iniparib plus chemotherapy in metastatic triple-negative breast cancer. New Engl. J. Med. 2011, 364, 205–214, doi:10.1056/NEJMoa1011418.
- Maeda, H.; Khatami, M. Analyses of repeated failures in cancer therapy for solid tumors: Poor tumor-selective drug delivery, low therapeutic efficacy and unsustainable costs. Clin. Transl. Med. 2018, 7, 11, doi:10.1186/s40169-018-0185-6.
- Mateo, J.; Ong, M.; Tan, D.S.; Gonzalez, M.A.; de Bono, J.S. Appraising iniparib, the PARP inhibitor that never was—What must we learn? Nat. Rev. Clin. Oncol. 2013, 10, 688–696, doi:10.1038/nrclinonc.2013.177.
- Whittle, J.R.; Lewis, M.T.; Lindeman, G.J.; Visvader, J.E. Patient-derived xenograft models of breast cancer and their predictive power. Breast Cancer Res. BCR 2015, 17, 17, doi:10.1186/s13058-015-0523-1.
- Vagia, E.; Mahalingam, D.; Cristofanilli, M. The Landscape of Targeted Therapies in TNBC. Cancers 2020, 12, doi:10.3390/cancers12040916.
- NIH. The Cancer Genome Atlas Program. Available online: https://http://www.cancer.gov/about-nci/organization/ccg/research/structural-genomics/tcga (accessed on 24 April 2020).
- Holen, I.; Speirs, V.; Morrissey, B.; Blyth, K. In vivo models in breast cancer research: Progress, challenges and future directions. Dis. Models Mech. 2017, 10, 359–371, doi:10.1242/dmm.028274.
- Eccles, S.A.; Aboagye, E.O.; Ali, S.; Anderson, A.S.; Armes, J.; Berditchevski, F.; Blaydes, J.P.; Brennan, K.; Brown, N.J.; Bryant, H.E.; et al. Critical research gaps and translational priorities for the successful prevention and treatment of breast cancer. Breast Cancer Res. BCR 2013, 15, R92, doi:10.1186/bcr3493.
- Dai, X.; Cheng, H.; Bai, Z.; Li, J. Breast Cancer Cell Line Classification and Its Relevance with Breast Tumor Subtyping. J. Cancer 2017, 8, 3131–3141, doi:10.7150/jca.18457.
- Gisselsson, D.; Lichtenzstejn, D.; Kachko, P.; Karlsson, J.; Manor, E.; Mai, S. Clonal evolution through genetic bottlenecks and telomere attrition: Potential threats to in vitro data reproducibility. GenesChromosomes Cancer 2019, 58, 452–461, doi:10.1002/gcc.22685.
- Ben-David, U.; Siranosian, B.; Ha, G.; Tang, H.; Oren, Y.; Hinohara, K.; Strathdee, C.A.; Dempster, J.; Lyons, N.J.; Burns, R.; et al. Genetic and transcriptional evolution alters cancer cell line drug response. Nature 2018, 560, 325–330, doi:10.1038/s41586-018-0409-3.
- Fleming, J.M.; Miller, T.C.; Meyer, M.J.; Ginsburg, E.; Vonderhaar, B.K. Local regulation of human breast xenograft models. J. Cell. Physiol. 2010, 224, 795–806, doi:10.1002/jcp.22190.
- Fung, A.S.; Lee, C.; Yu, M.; Tannock, I.F. The effect of chemotherapeutic agents on tumor vasculature in subcutaneous and orthotopic human tumor xenografts. BMC Cancer 2015, 15, 112, doi:10.1186/s12885-015-1091-6.
- Cell Line-Derived Xenograft—CDX Model Studies in Mice. Available online: https://http://www.criver.com/products-services/discovery-services/pharmacology-studies/oncology-immuno-oncology-studies/oncology-models/cell-line-derived-xenograft-cdx-mouse-models?region=3696 (accessed on 29 June 2020).
- Cell-Line-Derived Xenograft Models. Available online: https://http://www.creative-animodel.com/animal-model-development/cell-line-derived-xenograft-models.html (accessed on 29 June 2020).
- DeRose, Y.S.; Wang, G.; Lin, Y.C.; Bernard, P.S.; Buys, S.S.; Ebbert, M.T.; Factor, R.; Matsen, C.; Milash, B.A.; Nelson, E.; et al. Tumor grafts derived from women with breast cancer authentically reflect tumor pathology, growth, metastasis and disease outcomes. Nat. Med. 2011, 17, 1514–1520, doi:10.1038/nm.2454.
- Hidalgo, M.; Amant, F.; Biankin, A.V.; Budinska, E.; Byrne, A.T.; Caldas, C.; Clarke, R.B.; de Jong, S.; Jonkers, J.; Maelandsmo, G.M.; et al. Patient-derived xenograft models: An emerging platform for translational cancer research. Cancer Discov. 2014, 4, 998–1013, doi:10.1158/2159-8290.CD-14-0001.
- Sulaiman, A.; Wang, L. Bridging the divide: Preclinical research discrepancies between triple-negative breast cancer cell lines and patient tumors. Oncotarget 2017, 8, 113269–113281, doi:10.18632/oncotarget.22916.
- Choi, Y.; Lee, S.; Kim, K.; Kim, S.H.; Chung, Y.J.; Lee, C. Studying cancer immunotherapy using patient-derived xenografts (PDXs) in humanized mice. Exp. Mol. Med. 2018, 50, 99, doi:10.1038/s12276-018-0115-0.
- Brehm, M.A.; Shultz, L.D.; Luban, J.; Greiner, D.L. Overcoming current limitations in humanized mouse research. J. Infect. Dis. 2013, 208 (Suppl 2), S125–S130, doi:10.1093/infdis/jit319.
- Garcia, S.; Freitas, A.A. Humanized mice: Current states and perspectives. Immunol. Lett. 2012, 146, 1–7, doi:10.1016/j.imlet.2012.03.009.
- Laudanski, K.; Stentz, M.; DiMeglio, M.; Furey, W.; Steinberg, T.; Patel, A. Potential Pitfalls of the Humanized Mice in Modeling Sepsis. Int. J. Inflamm. 2018, 2018, 6563454, doi:10.1155/2018/6563454.
- Yeadon, J. Immunodeficient Mice for Cancer Studies: Which Host Strain Should I Use? Available online: https://http://www.jax.org/news-and-insights/jax-blog/2013/july/which-host-strain-should-i-use (accessed on 29 June 2020).
- Eyre, R.; Alferez, D.G.; Spence, K.; Kamal, M.; Shaw, F.L.; Simoes, B.M.; Santiago-Gomez, A.; Sarmiento-Castro, A.; Bramley, M.; Absar, M.; et al. Patient-derived Mammosphere and Xenograft Tumour Initiation Correlates with Progression to Metastasis. J. Mammary Gland Biol. Neoplasia 2016, 21, 99–109, doi:10.1007/s10911-016-9361-8.
- STOCK Trp53tm1Brd Brca1tm1Aash Tg(LGB-cre)74Acl/J. Available online: https://http://www.jax.org/strain/012620 (accessed on 29 June 2020).
- Menezes, M.E.; Das, S.K.; Emdad, L.; Windle, J.J.; Wang, X.Y.; Sarkar, D.; Fisher, P.B. Genetically engineered mice as experimental tools to dissect the critical events in breast cancer. Adv. Cancer Res. 2014, 121, 331–382, doi:10.1016/B978-0-12-800249-0.00008-1.
- Dabydeen, S.A.; Furth, P.A. Genetically engineered ERalpha-positive breast cancer mouse models. Endocr. -Relat. Cancer 2014, 21, R195-208, doi:10.1530/ERC-13-0512.
- Greenow, K.R.; Smalley, M.J. Overview of Genetically Engineered Mouse Models of Breast Cancer Used in Translational Biology and Drug Development. Curr. Protoc. Pharmacol. 2015, 70, 14 36 11–14 36 14, doi:10.1002/0471141755.ph1436s70.
- Ben-David, U.; Ha, G.; Khadka, P.; Jin, X.; Wong, B.; Franke, L.; Golub, T.R. The landscape of chromosomal aberrations in breast cancer mouse models reveals driver-specific routes to tumorigenesis. Nat. Commun. 2016, 7, 12160, doi:10.1038/ncomms12160.
- Fry, E.A.; Taneja, P.; Inoue, K. Oncogenic and tumor-suppressive mouse models for breast cancer engaging HER2/neu. Int. J. Cancer 2017, 140, 495–503, doi:10.1002/ijc.30399.
- Sledge, G.W., Jr. Curing Metastatic Breast Cancer. J. Oncol. Pract. 2016, 12, 6–10, doi:10.1200/JOP.2015.008953.
- Estimates for Funding for Research for Metastatic Disease: LOW. Available online: http://mbcn.org/research-funding/(accessed on 12 May 2020).
- EC. CORDIS. Available online: https://cordis.europa.eu/projects/en (accessed on 23 April 2020).
- Milosevic, M.; Jankovic, D.; Milenkovic, A.; Stojanov, D. Early diagnosis and detection of breast cancer. Technol. Health Care Off. J. Eur. Soc. Eng. Med. 2018, 26, 729–759, doi:10.3233/THC-181277.
- Han, X.; Wang, J.; Sun, Y. Circulating Tumor DNA as Biomarkers for Cancer Detection. Genom. Proteom. Bioinform. 2017, 15, 59–72, doi:10.1016/j.gpb.2016.12.004.
- Rawla, P. Epidemiology of Prostate Cancer. World J. Oncol. 2019, 10, 63–89, doi:10.14740/wjon1191.
- Taitt, H.E. Global Trends and Prostate Cancer: A Review of Incidence, Detection, and Mortality as Influenced by Race, Ethnicity, and Geographic Location. Am. J. Men’s Health 2018, 12, 1807–1823, doi:10.1177/1557988318798279.
- Pienta, K.J.; Abate-Shen, C.; Agus, D.B.; Attar, R.M.; Chung, L.W.; Greenberg, N.M.; Hahn, W.C.; Isaacs, J.T.; Navone, N.M.; Peehl, D.M.; et al. The current state of preclinical prostate cancer animal models. Prostate 2008, 68, 629–639, doi:10.1002/pros.20726.
- Ittmann, M.; Huang, J.; Radaelli, E.; Martin, P.; Signoretti, S.; Sullivan, R.; Simons, B.W.; Ward, J.M.; Robinson, B.D.; Chu, G.C.; et al. Animal models of human prostate cancer: The consensus report of the New York meeting of the Mouse Models of Human Cancers Consortium Prostate Pathology Committee. Cancer Res. 2013, 73, 2718–2736, doi:10.1158/0008-5472.CAN-12-4213.
- Cho, H.; Herzka, T.; Zheng, W.; Qi, J.; Wilkinson, J.E.; Bradner, J.E.; Robinson, B.D.; Castillo-Martin, M.; Cordon-Cardo, C.; Trotman, L.C. RapidCaP, a novel GEM model for metastatic prostate cancer analysis and therapy, reveals myc as a driver of Pten-mutant metastasis. Cancer Discov. 2014, 4, 318–333, doi:10.1158/2159-8290.CD-13-0346.
- Hsieh, C.L.; Xie, Z.; Liu, Z.Y.; Green, J.E.; Martin, W.D.; Datta, M.W.; Yeung, F.; Pan, D.; Chung, L.W. A luciferase transgenic mouse model: Visualization of prostate development and its androgen responsiveness in live animals. J. Mol. Endocrinol. 2005, 35, 293–304, doi:10.1677/jme.1.01722.
- Aggarwal, S.; Ricklis, R.M.; Williams, S.A.; Denmeade, S.R. Comparative study of PSMA expression in the prostate of mouse, dog, monkey, and human. Prostate 2006, 66, 903–910, doi:10.1002/pros.20413.
- Valkenburg, K.C.; Pienta, K.J. Drug discovery in prostate cancer mouse models. Expert Opin. Drug Discov. 2015, 10, 1011–1024, doi:10.1517/17460441.2015.1052790.
- Wei, C.; Willis, R.A.; Tilton, B.R.; Looney, R.J.; Lord, E.M.; Barth, R.K.; Frelinger, J.G. Tissue-specific expression of the human prostate-specific antigen gene in transgenic mice: Implications for tolerance and immunotherapy. Proc. Natl. Acad. Sci. USA. 1997, 94, 6369–6374, doi:10.1073/pnas.94.12.6369.
- Bullock, L.P. Brief overview of selected aspects of testicular hormone action. Environ. Health Perspect. 1981, 38, 11–18, doi:10.1289/ehp.813811.
- Oliveira, D.S.; Dzinic, S.; Bonfil, A.I.; Saliganan, A.D.; Sheng, S.; Bonfil, R.D. The mouse prostate: A basic anatomical and histological guideline. Bosn. J. Basic Med Sci. 2016, 16, 8–13, doi:10.17305/bjbms.2016.917.
- Michiel Sedelaar, J.P.; Dalrymple, S.S.; Isaacs, J.T. Of mice and men—warning: Intact versus castrated adult male mice as xenograft hosts are equivalent to hypogonadal versus abiraterone treated aging human males, respectively. Prostate 2013, 73, 1316–1325, doi:10.1002/pros.22677.
- Jeet, V.; Russell, P.J.; Khatri, A. Modeling prostate cancer: A perspective on transgenic mouse models. Cancer Metastasis Rev. 2010, 29, 123–142, doi:10.1007/s10555-010-9212-9.
- Kasper, S. Survey of genetically engineered mouse models for prostate cancer: Analyzing the molecular basis of prostate cancer development, progression, and metastasis. J. Cell. Biochem. 2005, 94, 279–297, doi:10.1002/jcb.20339.
- Kido, L.A.; de Almeida Lamas, C.; Marostica, M.R., Jr.; Cagnon, V.H.A. Transgenic Adenocarcinoma of the Mouse Prostate (TRAMP) model: A good alternative to study PCa progression and chemoprevention approaches. Life Sci. 2019, 217, 141–147, doi:10.1016/j.lfs.2018.12.002.
- Civenni, G.; Carbone, G.M.; Catapano, C.V. Overview of Genetically Engineered Mouse Models of Prostate Cancer and Their Applications in Drug Discovery. Curr. Protoc. Pharmacol. 2018, 81, e39, doi:10.1002/cpph.39.
- Irshad, S.; Abate-Shen, C. Modeling prostate cancer in mice: Something old, something new, something premalignant, something metastatic. Cancer Metastasis Rev. 2013, 32, 109–122, doi:10.1007/s10555-012-9409-1.
- Lin, D.; Wyatt, A.W.; Xue, H.; Wang, Y.; Dong, X.; Haegert, A.; Wu, R.; Brahmbhatt, S.; Mo, F.; Jong, L.; et al. High fidelity patient-derived xenografts for accelerating prostate cancer discovery and drug development. Cancer Res. 2014, 74, 1272–1283, doi:10.1158/0008-5472.CAN-13-2921-T.
- Hara, T.; Kouno, J.; Kaku, T.; Takeuchi, T.; Kusaka, M.; Tasaka, A.; Yamaoka, M. Effect of a novel 17,20-lyase inhibitor, orteronel (TAK-700), on androgen synthesis in male rats. J. Steroid Biochem. Mol. Biol. 2013, 134, 80–91, doi:10.1016/j.jsbmb.2012.10.020.
- Yamaoka, M.; Hara, T.; Hitaka, T.; Kaku, T.; Takeuchi, T.; Takahashi, J.; Asahi, S.; Miki, H.; Tasaka, A.; Kusaka, M. Orteronel (TAK-700), a novel non-steroidal 17,20-lyase inhibitor: Effects on steroid synthesis in human and monkey adrenal cells and serum steroid levels in cynomolgus monkeys. J. Steroid Biochem. Mol. Biol. 2012, 129, 115–128, doi:10.1016/j.jsbmb.2012.01.001.
- Van Hook, K.; Huang, T.; Alumkal, J.J. Orteronel for the treatment of prostate cancer. Future Oncol. 2014, 10, 803–811, doi:10.2217/fon.14.35.
- NIH. Study to Investigate the Effects of Orteronel on the QT/QTc Interval in Patients with Metastatic Castration-Resistant Prostate Cancer. Available online: https://clinicaltrials.gov/ct2/show/results/NCT01549951?view=results (accessed on 23 April 2020).
- NIH. Study Comparing Orteronel Plus Prednisone in Participants with Metastatic Castration-Resistant Prostate Cancer. Available online: https://clinicaltrials.gov/ct2/show/results/NCT01193257 (accessed on 23 April 2020).
- Loddick, S.A.; Ross, S.J.; Thomason, A.G.; Robinson, D.M.; Walker, G.E.; Dunkley, T.P.; Brave, S.R.; Broadbent, N.; Stratton, N.C.; Trueman, D.; et al. AZD3514: A small molecule that modulates androgen receptor signaling and function in vitro and in vivo. Mol. Cancer Ther. 2013, 12, 1715–1727, doi:10.1158/1535-7163.MCT-12-1174.
- Wadosky, K.M.; Koochekpour, S. Therapeutic Rationales, Progresses, Failures, and Future Directions for Advanced Prostate Cancer. Int. J. Biol. Sci. 2016, 12, 409–426, doi:10.7150/ijbs.14090.
- Suzman, D.L.; Antonarakis, E.S. Castration-resistant prostate cancer: Latest evidence and therapeutic implications. Ther. Adv. Med Oncol. 2014, 6, 167–179, doi:10.1177/1758834014529176.
- NIH. Drugs Approved for Prostate Cancer. Available online: https://http://www.cancer.gov/about-cancer/treatment/drugs/prostate (accessed on 23 April 2020).