Bast fiber plants require a post-harvest process to yield useable natural cellulosic fibers, denoted as retting or degumming. It encompasses the degradation of the cell wall’s non-cellulosic gummy substances (NCGs), facilitating fibers separations, setting the fiber’s quality, and determining downstream usages. Due to the inconvenience of traditional retting practices, bacterial inoculum and enzyme applications for retting gained attention. Therefore, concurrent changes of agroclimatic and socioeconomic conditions, the conventional water retting confront multiple difficulties, bast industries become vulnerable, and bacterial agents mediated augmented bio-retting processes trying to adapt to sustainability. However, this process’s success demands a delicate balance among substrates and retting-related biotic and abiotic factors. These critical factors were coupled to degrade bast fibers NCGs in bacterial retting while holistically disregarded in basic research. In this study, a set of factors were defined that critically regulates the process and requires to be comprehended to achieve optimum retting without failure.
Note: The entry will be online only after author check and submit it.
1. Introduction
Natural fibers as raw materials have gained momentum, attributed to their unique characteristics in numerous biocompatible commodities preparations. These fibers surmounted hazardous petroleum-based materials, requiring optimum fiber quality and those quality parameters inclined by growing conditions, retting, and other processing [
1,
2,
3]. The raw fiber applications in numerous downstream uses differ concerning fiber quality consistency, i.e., chemical, or mechanical properties, mostly from retting [
4,
5].
It is the most intricate and obligatory process for producing natural cellulosic fiber from heterogeneous bast fiber plants. Along with the separation feasibility of cellulosic bast fibers from the intact binding of pants cell wall matrix through the elimination of non-cellulosic gummy substances (NCGs), increased favorable mechanical properties eventually validated the multiple value-added green commodities preparations. It improves the cellulose composition of fibers. Elimination of NCGs results in disaggregating the pectin–lignin matrix bounds the elementary fibers and produces fiber bundles. It enhances the wettability of fibers and their compatibility with the polar functional groups of the polymeric matrix, improving the adhesion to the polymer matrix, forming a more robust interface and therefore improved composite accomplishments [
6]. The most prominent drawback of using natural cellulose fiber in biocomposites preparation is its inconsistent fiber quality, mostly inclined from the retting process. An effective bio-retting method encompasses the maximum elimination of NCGs within the least retting period. Subsequently, an effective degumming process could produce the desired cellulose fibers necessary to utilize fiber concerning economic aspects and fiber quality [
7,
8,
9]. Unprocessed fibers carry admixtures of NCGs, which increased volumetrically thick fiber bundles. This fraction did not accept the loads transferred across the fiber–matrix interface uniformly cross section, thus determining a reduction in the effective fiber volume fraction [
10]. Besides these, the targeted biocomposites preparations such as sound absorption insulation materials, automobile, and aeronautical interior decorative materials, virgin-grade packaging materials achieved after effective elimination of NCGs through the retting process [
11,
12].
Existing retting approaches confront inconsistent fiber quality, pollution, and input deficit issues, consequently driving to search for alternative retting processes. However, the proliferation of variable quality attributes from under retting and over retting leads to internal fiber rot with low strength and remains gummy substances unblemished, respectively [
13,
14]. More extended degumming periods (14–28 days) carry a risk of damaging cellulose and yield lower fiber mechanical properties. The dispersion of retting effluents such as galacturonic acid, butyric acid, acetic acid, lactic acid, Ca
2+, Mg
2+, bicarbonate content, and aquatic ecosystem alters the aquatic flora and fauna [
15,
16,
17,
18]. Because of worldwide climatic changes, low or lopsided precipitation crushes the accessibility of vast water sources. These disadvantages are unequivocally interlinking with the fiber quality, cultivator’s benefits, and generally, the bast fiber industry’s endurance.
Innovative retting approaches, such as chemical, mechanical, osmotic, ultrasound, and microwave, were attempting to yield fiber; notwithstanding, every alternative has its impediments concerning pollution and significant expense [
19,
20,
21]. The microbial solutions, for example, commencement of expected bacterial agents, either live strains or enzymes in the retting cycle, have demonstrated a plausible procedure yet request some basic conditions for their ideal initiation. Among different retting components, for example, bast plants, bacteria, solvents, and related biotic and abiotic elements requires a fragile equilibrium to break up the intact binding of cell wall polysaccharides network for quality fiber recuperation. Choices of fitting bacterial strains dependent on their ideal enzymatic potentials, growth characteristics, gums qualities, and other related variables are examined here. The bacterial retting associated critical factors that are obligatorily required for attaining maximum biodegradation of bast fibers adhering NCGS were conferred here. Along with this typical bacterial augmented bio-retting process, its alternative practices of the process were also discussed. This finding may assist with finding appropriate elements for bast fiber bacterial retting measure
2. Bacterial Retting Process
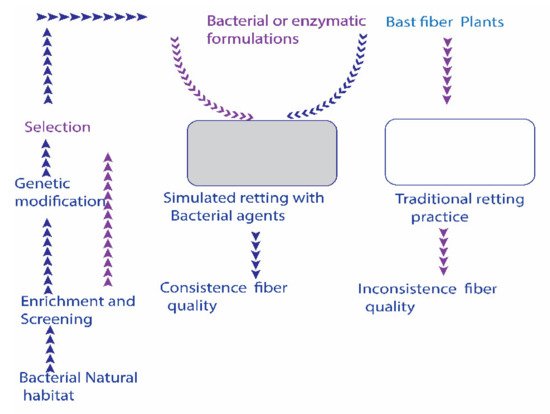
Utilizing bacterial retting agents is to inoculate specific bacterial strains or enzymes into the controlled or semi-controlled retting niches. Conversely, conventional retting happens by naturally developed microbiota in open field conditions. A schematic perspective on bacterial agents’ intervened retting measure is presented in Figure 1.
Figure 1. A possible scheme for incorporating bacterial bio-retting agents (enzymes + bacterial formulations) into non-sterile bast fiber retting niche. Inoculation of alkaline strains additionally surpasses the growth of unwanted endogenous bacteria. The establishment of a mechanized controlled retting niche could improve the retting process.
Macerating solvents, mainly water and bacterial extracellular enzymes combine to act on the insoluble plants’ surface. Bacterial enzymes initiate long-chain breaks in the cell wall gummy substances without the need to extricate these chains from their crystalline lattice. The significant strides of retting are water ingestion, an arrangement of calcium oxalate crystals; cortex expulsion; evacuation of gum in the center lamella and the fiber surfaces. The initial two cycles were finished under low enzyme activation, while the third cycle required maximum pectinase activity, and the last cycle commenced with higher pectinase and xylanase activities [
32,
33]. This retting process preserves the original carbohydrate structure in monomeric sugars, compared to the chemical destruction of the carbon, which possesses the upper limit of cellulose, however, a lesser lignin content [
34].
3. Bast Fiber Cell Wall Structure Influencing Bacterial Retting
Cellulosic bast fibers originate from a particular gelatinous kind secondary cell wall of dicotyledonous plants. The key features of this sort of cell wall are their high substance of cellulose accomplishing (85%), existing and contrasted with the classic xylan type cell wall [
35,
36,
37,
38]. Usually, the bast fibers cell wall average width is 4 to 10 µm; therefore, with the help of macerating solvents, bacteria successfully can break the cell wall barrier to invade [
39]. The elementary fibers are reinforced along with pectin’s and lignin outlining the following microstructure level, i.e., specialized technical fibers, with a distance across going from 50 to 100 μm [
40]. The primary cell wall is just 0.02 micrometers thick and made of lignin and pectin, and is a haphazardly situated cellulosic fiber, though the secondary cell wall is dominated by cellulose attached together with lignin and hemicellulose (
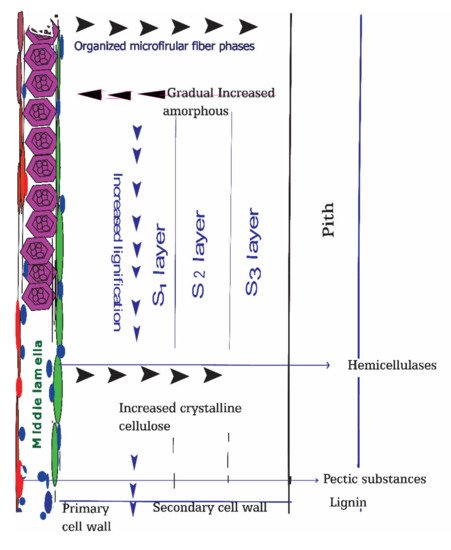
Figure 2).
Figure 2. Schematic representation of bast fiber plants cell walls and their compositions. Biodegradation of cell walls components at retting process is beneath their proportion of crystalline and amorphous structures and deposited gums concentrations such as degree of lignification and degree of pectins esterification.
The classical xylan-rich cell wall acts as the percussor of bast fibers gelatinous type cell wall [
41]. Due to hygroscopic nature of xylan, this type of a cell wall tends to degrade faster than those made of crystalline cellulose fiber. The presence of Rhamnogalacturonans-I in the gelatinous-type secondary cell walls effectively retained by cellulose microfibrils with separate layers termed as S1, S2, and S3 according to the order of their deposition. During the quick growth cycle of plant production, the most invasive noticeable output of primary bast fibers occurs [
42]. Fiber cell formations were categorized in two distinct phases: (a) expansion devoid of new stratum buildup called the primary cell wall and (b) step of cell wall setting designated as secondary walls [
43].
Cellulose accomplishes its most important focus in the S2 layer, and lignin accumulates in the center lamella and organized gathering in groups, isolated by the cortex parenchyma cell with the pectin and hemicellulose rich cell wall [
44]. Among three secondary cell wall layers, S2 is the more robust and determines the fiber properties. As cell wall structure and the bast fiber component’s orientation vary, the non-cellulosic gums in correlation with cellulosic fibers also vary in different positions. The outer, middle lamella is rich in pectic substances and lignin, which plays a crucial role in maceration and, subsequently, microbial invasion, and acts as an enzymatic epicenter. The organized fiber structure with crystalline compositions increases from the middle lamella to the secondary cell wall. Conversely, amorphous cell wall components were prevalent at outer, middle lamella and tended to decrease towards inner portions.
Different arrangement of crystalline in cellulose structure are brought about by intra and intermolecular hydrogen holding, which happened through hydroxyl bunches [
45]. There are 90–100 ‘g’ crystalline cellulose (Iβ)/100 g cellulose observed in plant-based fibers [
46]. Thus, the proportional orientation of bast components such as outer amorphous components degrade faster, and the inner crystalline cellulosic components take time to decompose. A proper level of retting within a minimum time is essential [
47] which gave the protection to the crystalline cellulosic fiber rather than amorphous gums.
4. Bast Fibers Cell Walls Constituents Affecting Retting Process
Bast fibers chemical constituents are categorized as major and minor groups. The main constituents for which the plants are domesticated are natural cellulose fiber, along with hemicellulose and lignin. On the other hand, pectic substances, ash, proteins, pigments, different extractives, and mineral constituents such as Ca and Mg. However, pectic materials present in trace quantities but determine the adhesive, mechanical, and defensive functions of bast fiber plants. Consequently, the separation of individual fiber through the retting process from a fiber bundle depends on the degradation of these pectic substances. The cellulose and lignin together shared for more than 60% and, at times, up to 80% of the secondary cell walls which also contain hemicelluloses and relatively less pectin compares to primary cell walls [
48,
49]. Major constituents of commercially important bast fibers are shown in
Table 1.
Table 1. Major cell wall compositions of commercially cultivated bast fiber plants (%).
| Bast Fiber |
Cellulose |
Hemicellulose |
Lignin |
Pectin |
Ref. |
| Jute |
59–72 |
12–20 |
12–13 |
2–4 |
[42] |
| Kenaf |
53.18 |
14.33 |
8.18 |
8.86 |
[43] |
| Hemp |
60–70 |
15–20 |
2–4 |
4 |
[44] |
| Ramie |
69–85 |
13–17 |
5–7 |
2 |
[40] |
| Flax |
62–72 |
17–21 |
2–5 |
2–3 |
[50] |
Cellulose is the repeating linear form of glucose unit composed of β-1, four linkages. Based on chemical composition, cellulose is the main component (50–99%) of all bast fibers. The percentage of cellulosic compositions differs from plant types and even varietal differences, determining the quality, and end use of fiber. Indication of retting reflects by removing its amorphous portions, which increases the crystallinity of that fiber. These single subunits together create a larger unit of microfibrils. The linkages throughout the microfibrils are closely interlinked, providing them with the excellent mechanical property. This hydrogen bond between the two cells with a mostly parallel cellulose layer is connected tightly and wrapped microfibrils. The microfibrils are then added to the crystalline cellulose fibers. Degradation of cellulosic components during retting destroys fibers. Crystalline cellulose portions could be beneficial for high-quality fiber; conversely, critical parts of cellulosic fragments gave lower fiber yield. Chemically, 17.5 percent of NaOH solutions are used to produce α-cellulose. The modern X-ray diffractometer technology determines the crystallinity of cellulosic fiber. Its proportion with other incrusted substances determines the fiber quality, mainly attributed to retting. The crystalline structure of cellulose protects it from degradation for a prolonged retting period.
The second abundant component of bast fiber is hemicellulose, which comprises complex branched groups of polysaccharide constituents with H-bonding to cellulose. These mixtures are sugar units such as xylan, xyloglucan, 4-methyl glucuronic acid, mannans, galacturonic acid, arabinogalactan, or other heteropolysaccharides residues. Galactoglucomamman is the major structural component of bast fibers hemicellulose. Differential characteristics of hemicellulose from cellulose include their degree of polymerization and lower weight. A higher proportion of hemicellulose conversely increases its water retention characteristics and made the fiber lower quality. The hemicellulosic cross-linking network considers the main structural constituent of the fiber cell. During the retting process, this fraction is not entirely removed by retting, and its proportion determines the crystallinity of cellulosic fiber. One of the significant functions of hemicellulase is to maintain the biodegradable characteristic of the fiber.
Pectin is the polymer of D-galacturonic acid residues originating from the middle lamella of the primary cell wall. Pectin is the chief incrusting material and the deciding factor for the retting, holding fiber bundles together with other cell wall components, ranging from 5 to 23 percent [
51,
52]. The middle lamella of bast fiber is the determinant hub of retting, where fiber cells are glued together with pectineus gums. This individual fiber makes the fiber bundles parallelly orientated to the stem’s longitudinal axis. Galactose composition is the typical gelatinous type of cell wall, also known as galactan and arabinogalactan. Pectin is one of nature’s most diverse biomacromolecules. In pectin structures, there are seventy different monosaccharide units, and seven different polysaccharides units observed. Pectineus substances differ from origins, which are inclined to esterified proportions (methylated) of total galacturonic unit contents. Its esterified variation demands specific pectinolytic enzymes to depolymerize [
53,
54].
Lignin consists of complex aliphatic and aromatic hydrocarbon polymers, and it generates powerful cellulose and hemicellulose clusters in plant materials. Lignification of bast fibers increases with the plants’ age and differs from sources, becoming more condensed than the hardwood fiber lignin [
55,
56,
57]. The hydrophobic lignin network distresses another network’s attributes so that it acts as a linking negotiator and surges the rigidity of the fiber to make composites materials [
58]. The full removal of lignin from the bast fiber cellulose portions during retting is a difficult task. Lignification in bast materials increases with the plants’ age and makes the fiber hard and whitish color. Conversely, delignification improves its color and reduces pore sizes, which is ideal for biocomposites materials. Another positive aspect of delignification is the reduced compositions of a hemicellulose-oriented acetyl group [
59].
A trace amount of wax, protein, soluble gums, and minerals components were also present on to cell walls of bast fibers, which could also play a role in the retting process. The most prominent roles play the cell walls calcium and magnesium minerals, combining with pectins forms Ca or Mg pectate. That pectate trapped the cellulose fiber on to the cell wall hard binding. Insoluble waxes are made of various alcohols and can extract with organic solutions.
5. Bacterial Strains Affecting Retting Process
All biological bast fiber retting processes are exclusively functions of microbial enzymatic activities. With the help of solvent maceration, bacteria penetrate the cell wall tissues through stomata by replacing intercellular air. The primary critical factors for choosing to ret potential bacteria depend on their optimal growth for specific nutrient requirements, temperature, pH, and O2 status. Spore-forming bacteria can survive in harsh environmental conditions and their extracellular enzymes are highly stable.
Plants endogenous bacterial mixture is non-selective, and their specific roles yet to be explored. In mixed microbial niche, some microbes secrete specific enzymes, and the rest of them areas non-producers, but all the microbes take nutrients from the sources for their metabolic activities. The non-enzyme producers are described as microbial cheaters and showed that higher costs of enzymes production favor cheaters and lower rate favored producers [
60]. The co-existence of cheaters and producers is highly organized but releases intermediate-level enzymes. Therefore, competition from non-selective cheaters is believed to result in inefficient enzyme production. On the other hand, nutritional limitations can restrict the microbial fostering and enzymatic degradation of heterogeneous lignocellulosic materials in natural atmospheres. The concept of bacteria applications in the retting process arises because of the tumbling efficiency of existing practices. Bacterial inoculum, whether natural selections or genetically modified strains, should reach the grower’s level as safe, be user-friendly, and have the potential to be stored for an extended period. Infrastructure related to bacterial retting was accomplishing with an artificial retting tank made of the cemented permanent tank, plastic tank, or temporary soil digging tank wrapping with plastic. Experimental retting tanks for fiber plants gave satisfactory results, both sterile and non-sterile retting at a research-level. The digestion of the lignocellulosic material by bacterial means in a controlled fermentation niche is a proven technology, however, it is still at the beginning phase for retting purposes.
Inoculation of these bacterial agents may serve to control the microbial diversity in the retting niche. Aerobic bacterial flora begins the retting process and continues if there prevails enough O2 for these bacteria’s sustenance. After that, anaerobic bacterial strains actively operate the retting process to the end. Till now, there was no evidence to use anaerobic stains as a starter inoculum. Facultatively anoxic strain actively participated in the retting process in both aerobic and anaerobic conditions. It is still debatable that a single isolate could complete the retting process or not, as multiple enzymes require retting completion. Application of bacterial consortia gave influential retting performances comparing to inoculation of single strains. Another possible solution could be the initiation of numerous enzymatic potentials strains. A list of retting potentials bacterial strains presents in Table 2.
Table 2. List of potentials bacterial strains uses for bast fiber retting.
| Bacteria |
Strain |
Enzymes |
Optimum pH |
Optimum Temp. |
Ref. |
| Bacillus sp. |
NT-39, 53, 76
MG-cp-2 |
PL, XL,
PG, Lac |
10 |
45 °C, 60 °C |
[61,62,63] |
| B. Pumilus |
NRRL B-212,
ATCC 7061 |
Exo-pectinase |
- |
- |
[64,65] |
| B. subtilis |
EFRL 01 |
PG |
8 |
45 °C |
[66] |
| B. licheniformis |
HDYM-03
KIBGE IB-21, SHG10, DSM-13 |
PL, PG, PNL |
- |
- |
[67,68,69,70] |
| B. Cereus |
HDYM-02 |
PG |
- |
- |
[71,72,73] |
| B. megaterium |
HDYM-09
AK2 |
PNL |
- |
- |
[67,74] |
| B. clausii |
- |
- |
- |
- |
[75] |
| B. tequilensis |
SV11-UV37
CAS-MEI-2-33 |
PL, PG
P |
10 |
40 °C |
[76,77] |
| B. halodurans |
M29 |
- |
|
- |
[78] |
| B. mojavensis |
M14, |
- |
- |
- |
[79] |
| Enterobacteriaceae Pectobacterium sp., Enterobacter lignolyticus |
DCE-01, SCF1 |
P, MN, XL
Lac |
- |
35 °C |
[80,81] |
Inoculation of bacteria strains decreases the other bacterial richness and diversity. The inoculated strain led to replacing different non-retting potentials bacterial strains; dominant groups are all degumming enzyme producers and survive until completion of retting. Application of pectinolytic bacteria as consortia gave superior retting results compared to single strains due to multiple enzymes requirement to degrade cell wall gummy substances [
32,
82,
83]. The main groups of bacteria that help to degrade lignin are Actinomycetes, Firmicutes, Bacteroides, and Proteobacteria [
84,
85,
86]. In several studies it was observe that the formulation of bacterial consortia in retting gave influential outcome comparing with using single strain. A list of bacterial consortia are presents in
Table 3.
Table 3. Bacterial consortia made for promoting bast fiber retting.
| Bacterial Consortia |
Composition |
Enzymatic Potentials |
Ref. |
| CRIJAF SONA |
Bacillus sp (PJRB1—Acc. No. MTCC 5573, PJRB2-MTCC 5574, and PJRB3-MTCC 5575) |
PG, PNL and Xylanase |
[83] |
| RAMCD407 |
P. aeruginosa, Enterococcus sp., B. subtilis, Bacillus sp. |
Pectinase, xylanase and mananase |
[32] |
| C-51, C-67, and C-90 |
B. megaterium, B. subtilis, B.s cereus, B. xiamenensis, B. koreensis, P. mirabilis, E. tabaci, K. oryzae, S. nematodiphila and Aeromonasjandaei. |
Pectinase and Xylanase |
[82] |
| MC1, MC2, MC3 |
B. subtilis B.s pumilus IMAU80221, B. pumilus GVC11 and B. pumilus SYBC-W |
Pectinase |
[16,20] |
6. Bacterial Retting Enzymes Affecting Retting Process
In the retting process, enzymes’ applications gave consistent fiber quality, restricted fiber damage, minimum or no fiber impurities, reduced odor, and retting pollution. Simultaneously, enzymatic breakdown of natural fibers matrix by pectinases, hemicellulase, and laccases is less invasive. Bast fiber biological retting is indeed the function of bacterial or fungal enzymes [
87]. Besides utilization of available commercial enzymes in retting could modify fiber characteristics as demanded properties that benefit bast fiber processing and fabric finishing ecology. Enzymatic separation produces stronger fiber suitable for textile applications [
88,
89,
90].
A group of diversified enzymes acts on pectin substances by cleaving the pectin chain’s 1,4-glycosidic bonds. So far, there have been records of 15 different types of pectinase. There have been several studies to investigate the effectiveness of pectinase on the bast fiber retting. As retting is the transformation of pectin [
87,
91], pectinases are the main retting enzyme. Based on its optimal pH activity, i.e., acid, and alkaline pectinases generally obtain from fungal and bacterial sources, respectively. Treating bast fibers with pectinase improves the mechanical characteristics of reinforced thermoplastic composites [
92,
93]. Alkaline pectinase has applications in the textile sector for bio-scouring and retting. Among alkali tolerant enzymes, polygalacturonase is the prime enzyme involved in the bast fiber retting process [
61,
94], including pectin lyase, pectate lyase, and esterase. When selecting pectinases, one should consider their esterification degree, as their polymerization and solubility depend on it. [
95]. Pectinase applications modify other non-cellulose plants’ gums, such as 3% pectinase application at kenaf retting reduced around one-half of the syringyl and guaiacyl lignin [
96]. Beyond this, the possible reason is pectin, and hemicellulase of bast materials restricts the same uronic acid content [
97]. Depolymerization reaction of pectic substances hangs on their chosen substrates, degradation mechanisms either trans-elimination or hydrolysis, cleavage mechanism either random for endo-acting or terminal for exo-acting enzymes [
98]. A list of retting pectinases and their mode of actins presents in
Table 4.
Table 4. List of bacterial enzymes involved in the biodegradation of non-cellulosic bast fibers gum in classical bacterial retting process.
| Name of Enzymes |
Enzyme Types |
Common Name |
E. C. Suggested
No. |
Substrate |
Cleavage Mechanism |
Product |
| Pectinases (Depolymerizes) |
Hydrolases |
Exo- polygalacturonases
(Exo-PG1 or PG |
3.2.1.67 |
Pectate |
Catalyze the α-1,4 glycosidic linkage |
mono galacturonate |
Exo- polygalacturonases
(Exo-PG2 or P.G.) |
3.2.1.82 |
Pectate |
Act on end cleaving site |
Di galacturonate’s |
| Lyases: (Polygalacturonate Lyase (PGL) |
Pectate lyases (Exo-PGL or P.L.) |
4.2.2.9 |
Pectin |
Act on end site bonds |
Unsaturated Di galacturonate’s |
Lyases: Polymethyl galacturonate Lyase
(PMGL) |
Pectin lyases
(Exo-PMGL or PNL) |
4.2.2.10 |
Pectin |
Random cleavage |
Unsaturated methyl oligo galacturonate |
Pectinases
(De-esterifying) |
Esterase |
Polymethyl galacturonate esterase (PMGE or PME) |
3.1.1.11 |
Pectin |
Random modification of ester group |
Pectin acid and methanol |
| Hemicellulose |
Hydrolases |
Xylanase |
3.1.1.8 |
Xylan
Mannan |
Cleavage β-1,4 glycosidic linkage |
xylose |
| Cellulase |
Hydrolases |
Endoglucanase |
3.2.1.4 |
Amorphous
Cellulose |
Nonreducing end of the cellulose chain |
glucose |
| Laccase |
Peroxidases |
Laccase
Mn peroxidase |
1.10.3.2
1.11.1.13 |
Lignin |
- |
p-quinone |
The production and application of pectinase from recombinant microbial sources have increased many folds over the last decades. A single-cell prokaryotic production of enzymes from genetically manipulated bacteria is always preferential due to their known genetic configuration. Whereas fungal sources pectinases are acidic and mainly uses in food sectors. The recombinant alkaline pectinases from
Bacillus clausi,
Penicilliums sp. 0602,
Bacillus halodurans, and
Bacillus lichiniformis were reported by several authors in the
E. coli expression system and observed their increased enzyme activity [
78,
99,
100].
Bacillus species prominently produce pectinase because of their alkali tolerance, and their cheap fermentation process. Alkaline
Bacillus species are salt-loving bacteria, such as
B. subtilis gave maximum pectate lyase activity by using ammonium salt as nitrogen sources from [
101]. Bacterial species belonging to the multiple enzymes encoding genes such as, polygalacturonase from
Bacillus licheniformis stain number KIBGE-IB21 contain other auxiliary enzymes such as pectin lyase and pectin methyl esterase with trace amounts of cellulase [
68].
Next to pectinases, hemicellulase is the second most important enzyme to contribute in the bast fiber retting process. Among different hemicellulases, xylanases are well characterized and act on the linear backbone of xylan. These hydrolytic enzymes degrade the lignocellulosic interphasic bast fibre xylan through retting process by cleaving β-1-4-D-xylosidic bond. A list of commercial enzymes use in bast fiber retting and processing shows in Table 5.
Table 5. List of commercial enzymes use in bast fiber retting and processing.
| Product |
Producers |
Activities |
| ScourzymeL |
Novozymes |
PG, PL |
| Bioprep 3000L |
Novozymes |
P |
| Texazym BFE |
Inotex |
P |
| Texazym SCW INOTEX Ltd., |
Inotex |
P, Xy, EG |
| Texazym DLG |
Inotex. |
Xy, CeB, EG, LPO |
| Texazym SER-3 |
Inotex |
P, Xy, EG |
| Texazym SER-4 |
inotex |
P, Xy, EG, CeB |
| Texazym SER-5 |
inotex |
P, Xy, EG, CeB and Lip |
| Baylase EVO |
Lanxess |
P |
| Lyvelin |
Lyven |
P |
Bast fiber xylan properties identify as arabinose-4-O-methylglucuronoxylan which typically occurs in softwood fibre plants. Usually, the xylanases are in diversified as like their substrate and works on specific substrates. Cellulase-free and alkaline resistant xylanase enzymes favor the retting process. In contrast to fungal origins, bacterial xylanase is alkali-tolerant, which is convenient for bast fiber retting.
Bacillus species are also well known for their alkali tolerant cellulase free xylanases [
62]. Other bacterial sources of xylanase reported from Clostridium
cellulovorans,
Erwinia spp.,
Bacillus halodurans,
Bacillus circulan.In general, cellulases for retting purposes consider as detrimental for the primary cellulosic bast fiber. The two different forms of cellulose, i.e., crystalline and amorphous, are degraded by differential cellulases. However, complete degradation of cellulose requires multiple cellulase enzymes. Usually, cellulose is insoluble and crystalline. Therefore, it tends to be nondegradable at a certain retting period whereas endoglucanases act on amorphous regions of the cellulose to degrade its glycosidic linkages’; the amorphous portions are not desired into the cellulosic fiber. In contrast, exo-glucanase or cellobiohydrolases acts on the crystalline portions of cellulosic bast fiber and strongly prohibited for retting purposes. A combination of cellobiohydrolases and endoglucanases entirely degrades the crystalline portions of cellulose [
102]. Scarification of this crystalline requires only a small mix of these enzymes. Among bacterial species, cellulase production is dominant in clostridium species with xylanase and or pectinases. Therefore, for retting purposes, cellulase-free xylanase and pectinase enzymes are preferential. A Clostridium strain C. cellulovorans genome study confirms its non-cellulolytic gene encoding hemicellulose and pectinase degrading enzymes [
103].
Laccases including phenol oxidase, laccase, and peroxidases are lignin degrading enzymes. There are three peroxidases identified involved in lignin degradation: lignin peroxidase, manganese peroxidase, and flexible peroxidase. As phenolic hydroxyl groups linked together by ether bonds in lignin molecules, useful lignin biodegrades must be broken down this to degrade lignin. In comparison to other microbial lignin sources, bacterial lignin has a high temperature tolerance in salty, alkaline environments [
104], which also favor pectin degradation. A new class of lignin peroxides known as dye-decolorizing peroxides was prominent in bacteria assumed have potentiality in the retting process. There are three classes of ligninolytic bacteria identified, i.e., actinobacteria, α-proteobacteria, and γ-proteobacteria, prevalently harbors in the aquatic ecosystem [
105]. Laccases are oxidases with a wide range of aromatic compounds such as phenolic, amine hydroxy idols, and benzenethiols. Bacterial sources of Laccases are reported from
Bacillus subtilis,
Streptomyces lavendulae, and
Sinorhizobium meliloti. Lignin peroxidase non-specifically degrades its polymers and is known as true laccase for its superior redox potentiality. The bacterial sources lignin peroxidases are abundant in actinobacteria and streptomyces. Manganese-dependent peroxidases were predominantly found in
Bacillus and
Panebacillilus species.
7. Interaction of Others Retting Factors
The bast plant’s age has a pivotal role in bacterial retting, as its composition changes at different growth stages. Over matured bast plants have gone through a coarse lignification process; therefore, lignin’s composition becomes denser. Comparing with the elongated portion, the base of the plants got more lignin than other parts. Physiochemical properties bast plants with ages have changed, such as fiber diameter increases with maturity, but primary cellulose content and moisture decrease. The other non-cellulosic components might be increases, and water reduction made it hard to swell up while enzymatic degradation [
106]. The harvesting period of jute plants for optimum retting and fiber quality between 110 to 120 DAS; overmature plants take a more extended period and yield low-quality fiber [
107]. Exposed to a long day photoperiod during growing season, bast fiber crops attain faster growth before flowering. Therefore, the green stem at vegetative maturity tends to ret faster and continues up to flower initiation. Harvest time toward the start of seed development prompted easier decortication with no recognizable effect on the fibers’ strength. [
108]. Therefore, the critical harvest time for optimum bio-retting of bast plants at flowering or before flowering stages, long before the seed maturity [
109,
110,
111]. Harvesting at this stage also beneficial for products damping characteristics, which happened due to increased lignin concentration [
112]. The lignin and pectin content relies on their age and maturity, about half that of the cores of roots, and the same level as in the matured samples plants more than 50 days old plants fiber [
113,
114].
The biodegradation cycle of bast fiber gummy segments influenced by the temperature as it straightforwardly influences the microbial development and metabolic action of proteins. Most of the retting potentials bacteria are optimally grown in between 25 to 35 °C, which ideal for bacterial water retting helps to maintain fiber quality [
115]. However, in temperate regions, the field retting process lingers more than two months due to a lack of optimal microbial growth conditions.
Uses of alkaliphiles in retting are advantageous due to their adaptive mechanisms and extracellular stable enzymes in harsh environmental conditions. They are incredibly functioning in these punitive retting environments by maintaining a cytoplasmic pH for their adaptions between the gradient of the medium. Interestingly, alkaliphiles’ intracellular enzymes are like non-alkaliphiles [
116,
117]. At the beginning, pH of the retting media decreases as low as 4.6–4.9; when the acid dispersion from the biochemical conversion of cell wall materials ended, the retting media again tended to increase the pH. This critical point indicates retting completion after that expects over retting [
118]. Commercial enzymes and chelators mediated retting were unable to sequestering Ca from fiber parts at lower pH but higher pH confirm successful retting [
119]. In bio retting calcium particles unequivocally hinder plant tissue mellowing at acid pH, yet not in the non-enzymatic acid hydrolysis [
120].
Chelating agents are substances that respond with metal particles to shape a steady water-solvent complex. Cell wall minerals components such as Ca and Mg make their corresponding Ca or Mg pectate through the “egg box” structure. Therefore, applications of Ca sequestering agents such as EDTA, oxalic acid, phosphorus, and nitrogenous compounds as a chelator in the retting niche, advances the retting process. Cell wall’s minerals reacts with chelating agents such as EDTA, oxalic acid, phosphorous, and nitrogen and makes corresponding water-soluble complexes. Optimum activation of pectinase with 18 mM EDTA gave superior fiber quality comparing with no EDTA treatment [
121]. Alkali tolerant pectinases play a crucial role in retting by preventing contamination, permits to adopt an open retting system [
122,
123,
124].
This entry is adapted from the peer-reviewed paper 10.3390/fib9080052